Toxoplasmose - Définition
La liste des auteurs de cet article est disponible ici.
Cycle du toxoplasme et pathogénie
Les trois formes parasitaires

T. gondii ne peut se multiplier de manière sexuée que chez les Félidés, qui constituent ainsi ses hôtes définitifs, bien qu’il puisse infecter tous les animaux homéothermes, dénommés hôtes intermédiaires. Le toxoplasme a un cycle complexe qui implique la transmission entre hôtes par des stades spécialisés pour l’invasion (voir figure) :
- le stade tachyzoïte, forme proliférative infectieuse chez l’hôte intermédiaire, se développe dans des vacuoles transitoires qui peuvent contenir jusqu’à 128 parasites ; cette forme peut se retrouver aussi chez le fœtus.
- le stade bradyzoïte, chez l’hôte intermédiaire, est contenu dans des kystes intracellulaires qui mesurent environ 100 µm de diamètre et contiennent plusieurs milliers de parasites ;
- le stade mérozoïte, chez l’hôte définitif, est le seul stade capable de reproduction sexuée ;
- le stade sporozoïte, résultat de la reproduction sexuée chez l’hôte définitif, est libéré dans l’environnement avec les déjections du chat dans des oocystes de 10 à 15 µm de diamètre qui contiennent 8 sporozoïtes.
Les stades sporozoïte et bradyzoïte correspondent à des formes de résistance et de dissémination du parasite car les kystes et les oocystes protègent dans une certaine mesure, les parasites qu’ils contiennent des variations de température, de pH, etc. T. gondii peut alterner entre ces stades en fonction de son hôte et de son contexte, par un processus de différenciation.
- Forme végétative
- Le tachyzoïte ou trophozoïte : C’est la forme que prend le parasite seul. il est alors très fragile ; sa présence est toujours endocellulaire (il ne résiste ni à l’eau de Javel ni à l’acide chlorhydrique gastrique). L’ingestion n’est donc pas contaminante.
- Il se reproduit rapidement par un processus de multiplication asexuée (endodyogénie) chez l’hôte intermédiaire, toujours dans des macrophages. Puis il en sort en en perforant la paroi au moyen d’une protéine qu’il produit (perforine). Des parasites génétiquement modifiés pour ne pas produire cette protéine ne peuvent sortir du macrophage qui gonfle en formant une boule. Visuellement, l’enveloppe du parasite a la forme d’une goutte d’eau un peu arquée (toxon en grec signifie arc), d’environ 6 à 8 µm de longueur et de 3 à 4 µm de largeur. Le pôle postérieur arrondi contient le noyau tandis que le pôle antérieur plus aigu possède des ultrastructures adaptées à la pénétration cellulaire (complexe apical).
- Forme kystique
- Cette forme est plus résistante que la précédente (forme de résistance et de dissémination), entourée par une membrane épaisse, de forme sphérique ou ovoïde, elle mesure de 50 à 200 µm. Elle contient en plusieurs milliers d’exemplaires une forme végétative particulière le bradyzoïte ou cystozoïte (3 à 4 microns), un kyste de 100 µm en contient 2 000 à 3 000. Les bradyzoïtes résultent d’une série de multiplications asexuées, colonisant l’intérieur d’une cellule hôte. Leur multiplication est assez lente, et ne peut se faire que dans une cellule nerveuse ou musculaire de l’hôte intermédiaire. Dans les tissus, les kystes restent longtemps vivants, produisant des antigènes qui entretiennent l’immunité. Les kystes peuvent survivre plusieurs jours à température ambiante et plusieurs mois à 4 °C. Ils sont détruits par la chaleur (un quart d’heure à 56 °C) ou la congélation (24 heures à -20 °C).
- L’oocyste
- L’oocyste coccidien est très résistant, même à l’eau de Javel (forme de résistance et de dissémination), c’est la forme que l’on retrouve dans le milieu extérieur (sol, plantes…) où il effectue sa maturation en quelques jours (de un à cinq) à température ambiante et en présence d’oxygène. Sa résistance lui permet de rester vivant pendant plusieurs mois dans le sol, mais il est détruit par la chaleur lors de la cuisson, la dessiccation ou la congélation (-30 °C). Il est le résultat de la reproduction sexuée du parasite chez le chat. C’est un ovoïde de 15 µm par 10 µm regroupant 2 sporocystes de 6 à 8 µm de diamètre, contenant 4 sporozoïtes chacun (un sporozoïte ressemble à un tachyzoïte).
Le cycle de reproduction du parasite
Le cycle peut être direct, c’est-à-dire sans hôte intermédiaire (cycle monoxène ou court) ou indirect en passant par un ou plusieurs hôtes intermédiaires (cycle hétéroxène ou long).
L’hôte définitif du parasite est principalement le chat, mais les autres félidés sont aussi concernés.
Les hôtes intermédiaires sont tous les animaux à sang chaud : mammifères et oiseaux (le chat, hôte définitif se contamine en dévorant des oiseaux ou des souris, petits animaux qu’il chasse avec assiduité, lorsqu’il peut sortir à l’extérieur).
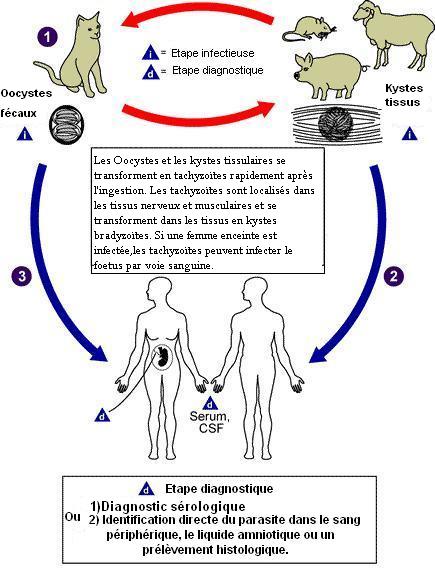
Les membres de la famille des chats (Felidae) sont les seuls hôtes définitifs connus pour les étapes sexuées de T. gondii et représente le principal réservoir de l'infection. Les chats sont d'abord infectés (étape 1 de la figure) en mangeant de la viande contenant des formes kystiques de T.G ce qui aboutit à la formation, dans son intestin, de gamétocytes dont la fusion donne des oocystes, éliminés dans les selles. Ceux-ci peuvent survivre dans le milieu extérieur, où ils se transforment en sporocystes puis en sporozoïtes infectants, qui sont ingérés par des animaux tels que des rongeurs, des moutons ou des porcs. Le parasite quitte le tractus digestif et, au stade trophozoïte, gagne divers tissus, se multipliant dans les macrophages et survit dans les muscles, le cœur, le cerveau sous formes de kystes contenant les bradyzoïtes, ce qui entretient l'immunité du sujet infecté. L'humain peut être infecté de différentes façons : A) ingestion de viandes crues infectées contenant des formes kystiques de T.G (étape 2 de la figure) ; B) ingestion d'oocystes provenant de fèces de chat à partir de mains ou de viandes contaminées (étape 3 de la figure) ; C) transplantation d'organe ou transfusions sanguine ; D) transmission congénitale de la mère au fœtus ; E) inoculation accidentelle de tachyzoites.
Contamination initiale
Les oocystes sont présents sur les plantes ou la terre souillées par des déjections d’animaux (chats en particulier). De là, ils peuvent contaminer les aliments, les mains ou l’eau de boisson, puis être ingérés.
La présence des kystes dans la viande est fréquente : 80 % des ovins et des caprins adultes sont contaminés, le porc est généralement contaminé dans moins de 40 % des cas, les autres espèces animales peuvent toutes être contaminées mais dans des proportions inconnues.
Lorsque la viande est consommée crue ou insuffisamment cuite, les kystes ne sont pas détruits et s’installent dans l’organisme hôte. Les kystes ne passent pas la barrière placentaire, seuls les trophozoïtes ou tachyzoïtes passent, si la maladie se déclenche en cours de grossesse, et des cas de contamination par greffe d’organe ou transfusion sanguine ont été recensés.
Reproduction
Hôte intermédiaire
Chez l’hôte intermédiaire, les oocystes libèrent les sporozoïtes, lesquels libèrent les tachyzoïtes (ou trophozoïte) au niveau du tube digestif, et vont passer la barrière intestinale. Ils vont se reproduire dans les cellules de l'hôte (la pénétration se fait par un mécanisme actif et non par phagocytose), déclenchant une phase sanguine de dissémination ou septicémie : l’hôte développe la toxoplasmose.
La réponse immunitaire de l’hôte confine ensuite le parasite à l’intérieur des organes dans lesquels la réponse immunitaire est la plus faible : l’œil, le cerveau, les muscles. Les parasites s’y enkystent, les kystes contiennent de nombreux bradyzoïtes et sont en attente d’une éventuelle réactivation. Cette réactivation se produit lorsque les chairs contenant des kystes sont consommées par un nouvel hôte n’ayant pas encore développé la maladie ou immunodéprimé, ou lors d’une greffe d’organes contenant ces kystes ; la réactivation de bradyzoïtes en tachyzoïtes a été décrite chez l'animal, ce qui permet de comprendre en partie ce qui se passe chez l'immunodéprimé, mais les mécanismes exacts demeurent toujours inexpliqués chez ce type de patient.
Hôte définitif
Chez l’hôte définitif, le parasite ingéré (généralement en dévorant un rongeur ou un oiseau infecté) se localise dans le tube digestif, provoquant une coccidiose. Le parasite produit alors des oocystes par reproduction asexuée puis sexuée ; en effet, les trophozoïtes libérés se multiplient au niveau du tube digestif. Il va se produire une reproduction sexuée avec formation de microgamètes mâles et de macrogamètes femelles ; la fécondation conduit aux oocystes. Ces oocystes seront rejetés dans l’environnement de l’hôte avec ses déjections, mais les excréments ne sont généralement pas contaminants pendant les deux premiers jours qui suivent l'excrétion. Les oocystes nécessitent une maturation de 14 jours pour devenir potentiellement pathogènes et résistent environ 1 an dans le milieu extérieur. Chez le chat par exemple, environ 2 % d’entre eux disséminent des oocystes, sur des périodes allant de une à trois semaines. Des études montrent qu’ensuite l’élimination ne se reproduit pas, même après de nouvelles expositions au parasite. Ce sont donc, en général, les jeunes chats qui excrètent le parasite. Bien que l'agent pathogène ait été détecté sur la fourrure des chats, il n'a pas été retrouvé sous une forme infectieuse, et une infection directe consécutive à la manipulation des chats est généralement considérée comme très rare.