Sarcophagidae - Définition
La liste des auteurs de cet article est disponible ici.
Biologie des Sarcophagidae
Les Sarcophagidés sont larvipares ou ovilarvipares, c’est-à-dire ils déposent larves ou œufs dont, immédiatement, éclosent les larves. Ce fait montre que les larves passent un certain temps dans l’organisme maternel, constituant ce qui s’appelle le parasitisme stationnaire. Un cas particulier est représenté par le genre Helicobosca (ou Eurychaeta) dont le développement larvaire a lieu dans l’abdomen maternel jusqu’au deuxième âge ou même quand il est nécessaire la nymphose.
Endoparasites des Lépidoptères
On sait maintenant avec certitude que les Sarcophagidés parasitent un nombre plus ou moins grand d'espèces de papillons. La difficulté de l’identification de ces diptères fait que ce phénomène est complètement ignoré. Autrefois, de la culture de Porthetris dispar L de Roumanie ont été obtenues Liosarcophaga portschinskyi Rohdendorf et des élevages du même hôte de Sardaigne, dans les années 1972-1977, ont été identifiées quelques espèces, parmi lesquelles: Liosarcophaga exuberans (Pandellé), Liosarcophaga tuberosa (Pandellé), Varirosellea uliginosa (Kramer), Liosarcophaga portschinskyi Rohdendorf, Curranea tibialis (Macquart), Sarcotachinella affinis (Fallen) et Thyrsocnema corsicana (Villeneuve). Excepté Sarcotachinella affinis (Fallen), qui est connue comme parasite de plusieurs hôtes de Geometrides, Lymantriides, Hyponomeutides et Lasiocampides, les autres espèces ont été mentionnées pour la première fois comme entomophage (Lehrer & Luciano, 1980).
Quelques espèces sont polyphages, ayant plusieurs hôtes. Ainsi, pour l’espèce nord-américaine Sarcodexia sternodontis Townsend on mentionne presque trente espèces de papillons.
Endoparasites des Orthoptères
Certains Sarcophagides parasitent les Orthoptères même au moment de leur éclosion. Les genres Blaesoxipha Loew, Sarcophila Rondani etc. attaquent les Grylides et d’autres genres d’Acridides, qui les poursuivent en vol sur une distance de plusieurs kilomètres.
Ils peuvent être univoltins ou plurivoltins, d’après le nombre des générations qui se succèdent dans un an. Blaesoxiphella brevicornis Villeneuve a trois générations par an et Acridophaga caridei (Brethes) 7-8 générations par an. La durée de chaque génération varie entre 1 et 2 mois. Chez Blaesoxipha krameri Baranov, la durée d’une génération est d’un mois, chez Acridophaga caridei (Brethes) elle est de 32-64 jours et davantage chez quelques espèces américaines.
L’accouplement peut avoir lieu 24 heures après l'éclosion de l’imago ou 8-12 jours après la maturité sexuelle et peut durer de 15 minutes (Blaesoxipha filipjevi Rohdendorf) jusqu’à 2 heures [(Blaesoxipha laticornis (Meigen)] ou plus.
Les femelles peuvent éjecter 1 à 5 larves en une fois et cela de 1 à 6 fois par jour, jusqu’à un total de 180 à 400 larves.
Les femelles de Blaesoxipha filipjevi Rohdendorf, Blaesoxipha unicolor (Villeneuve) ou Acridophaga caridei (Brethes) poursuivent les Acridiens du sol, tandis que Blaesoxipha krameri Baranov ceux pendant le vol. Les deux premières espèces prjettent les larves à distance sur leurs hôtes ; la dernière les dépose sur leur corps.
Dans le cas des espèces avec ovipositeur, les femelles habituent les hôtes à leur présence, par manœuvres de rapprochement, puis déposent les pattes sur eux et introduisent brusquement une ou plusieurs larves dans une région déterminée du corps: Blaesoxipha laticornis (Meigen) et Blaesoxipha berolinensis Villeneuve dans la cavité génito-anale; Blaesoxipha ungulata (Pandellé) dans la cavité buccale; Blaesoxiphella brevicornis Villeneuve et Servaisia rossica (Villeneuve) dans les plis intersegmentaux des tergites abdominaux; Protodexia hunteri (Hough) à la base des pattes; Servaisia falciformis (Aldrich) inocule les larves par la cuticule des metafémurs de l'hôte.
Dans les hôtes les larves sont plasmophages, mais à la fin de la période du développement elles deviennent sarcophages et représentent un véritable péril pour l’orthoptère parasité. La durée de la vie dans l’intérieur de l'hôte varie entre 5-10 jours, et la métamorphose entre 1-4 semaines.
Parasites des arachnides
En 1986, Cantrell a observe dans l’Est de l’Australie les Sarcophagidae des genres Parasarcophaga (s. lat.) et Baranovisca Lopes qui parasitent les cocons des arachnides.
Endoparasites des vers de terre
En 1947 James considère que Sarcophaga carnaria Boettcher est un élément habituel des myiases sur certains pâturages de l’Europe, comme en Angleterre, Italie et Sicile, où elle produit la myiase des plaies chez les moutons.
Portschinsky a exprimé l’opinion que ces cas de myiases ne sont pas déterminés par Sarcophaga carnaria Boettcher, mais par Wohlfahrtia magnifica (Schiner). Kirchberg, en 1954 a affirmé que la première espèce est aussi un parasite obligatoire des vers de terre.
Myiasigènes
Les diptères de cette famille ont tendance à déposer leurs larves sur les plaies préexistantes ou sur les muqueuses intactes de plusieurs espèces d’animaux homéothermes, y compris l’homme, provoquant des disturbations fonctionnelles et, parfois, même leur mort. Néanmoins, seulement certaines espèces du genre Wohlfahrtia Brauer & Bergenstamm sont des parasites obligatoires et producteurs de myiases, les autres étant des myiasigènes facultatifs ou secondaires, mais presque aussi dangereux.
Dans la catégorie des myiasigènes obligatoires, Wohlfahrtia magnifica (Schiner) qui provoque la parasitose connue sous le nom de wohlfahrtiose, est répandue dans la région paléarctique, W. virgil (Walker) dans la région néarctique et W. nuba (Wiedemann) dans les régions paléarctique et éthiopienne.
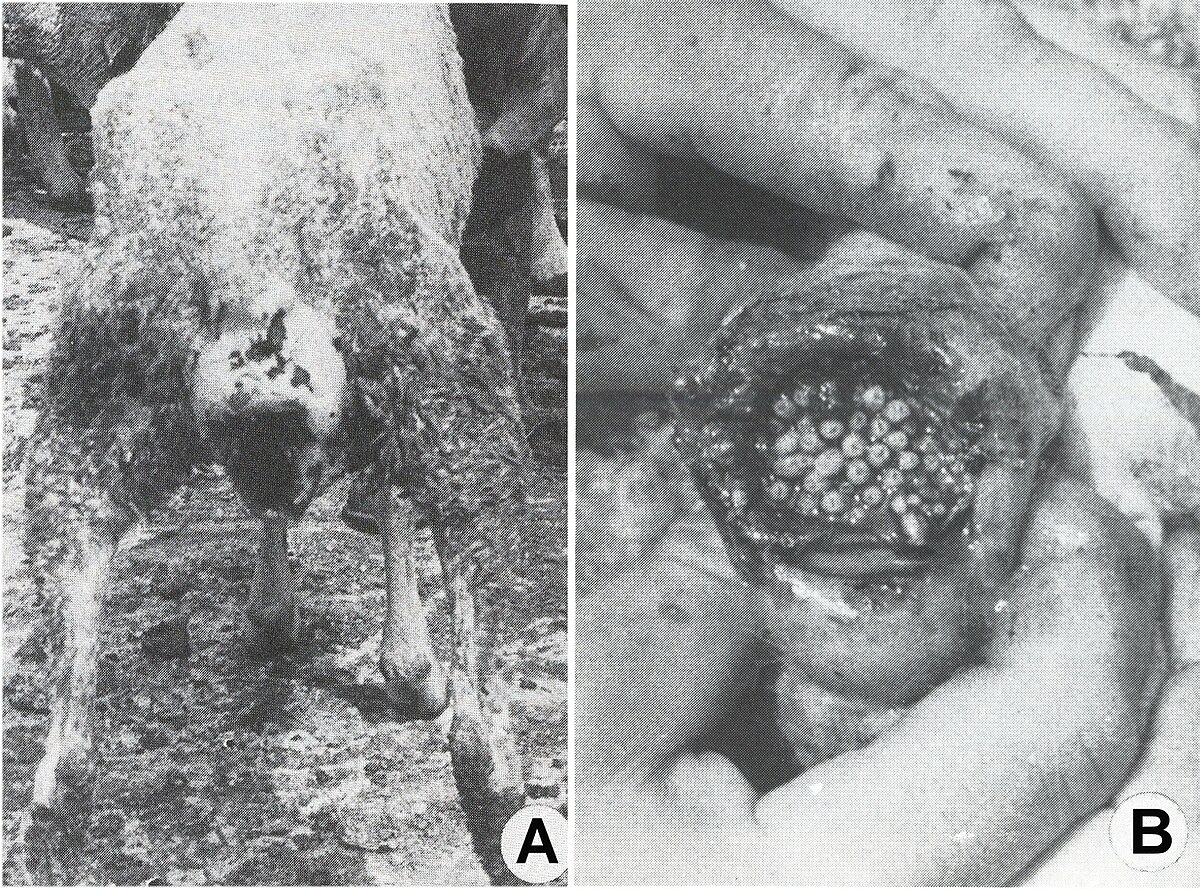
Wohlfahrtia magnifica attaque les moutons, les vaches, les chevaux, les chiens et d'autres animaux homéothermes, en déposant ses paquets successifs d'œufs et de larves en très grand nombre, sur les tissus humides ou les plaies de ceux-ci. Les larves s'enfoncent rapidement et profondément dans les tissus et, en quelques jours elles provoquent des lésions énormes, qui peuvent déterminer la mort des hôtes. Parfois, les hommes de garde des troupeaux d'animaux, qui ont des plaies infestées et pas traitées ou qui dorment à l'air libre, peuvent être infestés par ce sarcophage.
Pendant les recherches faites en Roumanie sur cette "wohlfahrtiose" (A. Lehrer & Fromunda, 1986; A. Lehrer, M. Lehrer & Verstraeten, 1988; A. Lehrer & Verstraeten, 1991), ont été observées des nombreuses myiases graves et très graves chez les moutons de race Mérinos ou Corrideli, importées d'Australie ou Nouvelle-Zélande, qui correspondent à certaines conditions d'élevage.
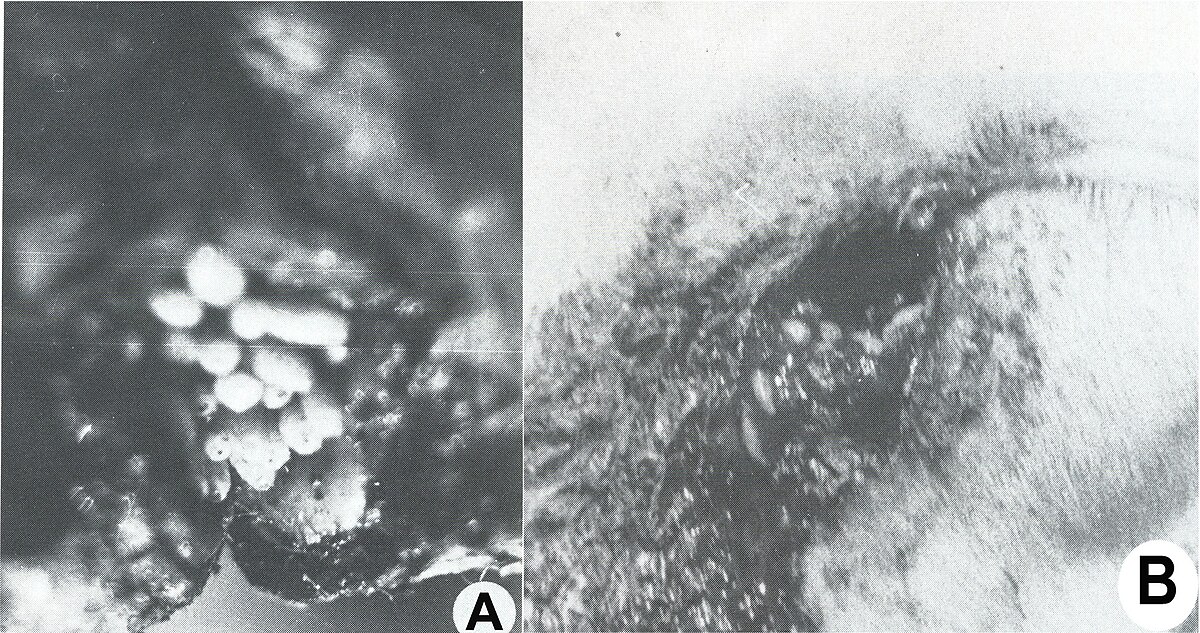
Après un court intervalle suivant leur importation, le parasite s'est multiplié vertigineusement et s'est répandu dans toutes les régions du pays, décimant 80-90% de l'effectif des moutons. La dispersion foudroyante du parasite est devenue un problème européen, parce que beaucoup de pays, qui ont voulu développer un secteur de moutons à laine fine et demi-fine, ont souffrit des préjudices économiques incalculables, surtout à la suite des myiases génitales : vulvaires (fig. 5 A) et du prépuce (fig. 5, B), des podomyiases (fig. 6, A) ou d’ophtalmomyiases (fig. 6, B), ainsi que des attaques mortelles des nouveau-nés.
De la catégorie des parasites facultatifs est identifié, plus ou moins correctement, un nombre de vingt-trois espèces, qui produisent notamment la myiase des plaies et des cavités naturelles, mais aussi celle intestinale des animaux et de l’homme. Ainsi, ont été mentionnées :
- Bercaea cruentata (Meigen) qui, ayant les larves coprophages très mobiles, peut provoquer tant la myiase rectale, que celle intestinale. Zumpt soutienne que les femelles déposent les larves pendant la défécation de l’homme et elles pénètrent rapidement dans le rectum par l’anus, pouvant compléter leur développement larvaire dans les intestins. Cette espèce produit aussi la myiase des plaies en Tripoli et dans l’île Maritaux.
- Boettcherisca peregrina (Robineau-Desvoidy) - infeste les plaies, étant accusée aussi de la myiase intestinale de l’homme dans l’Extrêmes Orient et les régions orientale et australienne.
- Ctenodasypygia fertoni Villeneuve - peut infester les plaies dans la région paléarctique.
- Curranea tibialis Macquart - a provoqué l’invasion des lésions de la tête, déterminées par Trichophytosis en Tripoli et, occasionnellement, la myiase intestinale de l’homme en Europe.
- Engelisca nodosa Engel - est accusée de myiase auriculaire et des plaies chez l’homme en Rhodésie.
- Jantia crassipalpis Macquart - est répandue dans les régions holarctique et australienne, étant mentionnée comme parasite secondaire des plaies aux moutons, en Serbie.
- Kellymyia plinthopyga Wiedemann - est mentionnée comme infestant les lapins et autres animaux en Guinée Britannique et Texas.
- Liopygia ruficornis Fabricius - produite la myiase cutanée en Inde aux l’homme, chiens, chevaux et bardeaux.
- Liosarcophaga babiyari Lehrer - les données parasitologiques de cette espèce ne sont pas sûres.
- Liosarcophaga ?misera Walker - Alwar & Seshian (1958) accuse cette espèce des myiases des plaies aux chameaux, bœufs et une vache de Madra. Mais, ces données ne sont pas sures parce que l’identification de l’espèce n’est pas correcte.
- Liosarcophaga tuberosa Pandellé - étant dans le groupe général “misera”, il n’y a aucune sûreté dans l’affirmation qu’elle provoque la myiase cutanée aux moutons.
- Parasarcophaga albiceps Meigen - mentionnée qu’elle a déterminé la myiase cutanée chez un taureau en Inde (Senior-White, Aubertin & Smart).
- Parasarcophaga hirtipes Wiedemann - très répandue en Extrême Orient et dans la région orientale, étant connue comme producteur de la myiase intestinale chez l’homme en Égypte. Elle détermine la myiase des plaies aux bétails et moutons en Rhodésie.
- Ravinia striata Fabricius - incriminée dans la myiase intestinale par Castellani & Chalmers (1919), mais par Trofimov, Tsuav & Alieva (1958) aussi en Russie.
- Sapromyia bullata Parker - accusée des myiases des plaies et intestinale chez l’homme dans les états du sud de la région néarctique.
- Sarcodexia sternodontis Townsend - provoque la myiase des plaies aux hommes et aux animaux dans le sud de l’Amérique, aux bétails en Floride et la myiase auriculaire dans la zone du canal du Panama.
- Sarcophaga carnaria Boettcher - les données sur son statut de myiasigène ne sont pas sûres, parce qu’elle est considérée comme parasite obligatoire des vers de terre (Kirchberg, 1954).
- Thomsonea argyrostoma Robineau-Desvoidy - infeste les plaies de l’homme et des animaux en Serbie, en association avec Wohlfahrtia magnifica Schiner.
- Titanogrypa pedunculata Aldrich - détermine la myiase auriculaire et celle des tissus nécrosés de l’homme dans la région du canal de Panama.
- Tricholioproctia froggati Taylor - parasite secondaire des moutons en Australie tropicale et subtropicale.

















































