Sarcophagidae - Définition
La liste des auteurs de cet article est disponible ici.
Classification
La taxonomie de cette famille constitue un foyer de discussions et disputes, parfois très passionnés, entre les chercheurs ayant des conceptions conservatoires et ceux qui utilisent des moyens de recherche plus modernes.
Liste des genres
Selon NCBI :
- sous-famille Miltogramminae
- genre Metopia
- genre Miltogramma
- genre Pterella
- genre Sphenometopa
- genre Taxigramma
- sous-famille Paramacronychiinae
- genre Brachicoma
- genre Goniophyto
- genre Nyctia
- genre Sarcophila
- genre Wohlfahrtia
- sous-famille Sarcophaginae
- genre Argoravinia
- genre Blaesoxipha
- genre Boettcheria
- genre Chlorosarcophaga
- genre Dexosarcophaga
- genre Duckemyia
- genre Engelimyia
- genre Fletcherimyia
- genre Helicobia
- genre Metoposarcophaga
- genre Notochaeta
- genre Oxysarcodexia
- genre Peckia
- genre Peckiamyia
- genre Ravinia
- genre Sarcodexia
- genre Sarcofahrtiopsis
- genre Sarconesia
- genre Sarcophaga
- unclassified Sarcophaga
- genre Sinopiella
- genre Spirobolomyia
- genre Titanogrypa
- genre Tricharaea
- unclassified Sarcophagidae
- genre Villegasia
Selon ITIS :
- genre Tephromyiopsis
- genre Udamopyga
- sous-famille Miltogramminae
- sous-famille Sarcophaginae
Taxonomie et classification d'après la morphologie des organes mâles
Acceptant que cette famille se partage en trois sous-familles (Miltogramminae, Paramacronychiinae et Sarcophaginae), bien que certaines intuitions taxonomiques prédéterminent encore d’autres groupes importants.
La plus importante sous-famille est Sarcophaginae, qui comprend un très grand nombre de taxons génériques et spécifiques et qui connaît un développement intéressant d’un seul genre initial Sarcophaga Meigen, 1824. Après une période d’accumulation initiale de données, plus ou moins sommaires et très généralisées, les études de G. Böttcher (1912-1913) ont constitué un moment crucial pour la taxonomie de la famille. Observant la grande convergence des caractères somatiques, il a abordé sous une manière scientifique, l’étude des genitalia mâles des espèces et a prouvé que leurs complexité et configuration sont les éléments les plus sûrs pour l’identification des taxons, de rang différent.
Boris B. von Rohdendorf (1937) est une des figures emblématiques de l'étude de la taxonomie moderne des Sarcophagides. Il montre que les esquisses des génitalia mâles présentées par Böttcher sont trop schématiques et que, par méthodes histologiques et graphiques plus évoluées, on peut découvrir de nombreux taxons inconnus. Par cette conception, il fixe les bases scientifiques du système d'étude taxonomique de la sous-famille Sarcophaginae, tant pour la région paléarctique, que pour celle éthiopienne (1963). Sa méthodologie a été suivie par un nombre important de chercheurs: Baranov, Cepelak, Lehrer, Povolny, Fan Zide, Lopes, Kano, Shinonaga, Mihalyi, Verves etc. Ils considèrent que le type de structure des génitalia mâles et, notamment, du phallosome, reflète mieux la valeur hiérarchique des unités taxonomiques fondamentales de genre et d’espèce, due à sa grande stabilité génétique. Cette vision s'oppose fermement à celle d'un autre diptérologiste français, Eugène Séguy (1941), qui a adopté l’unité générique Sarcophaga.
Après soixante ans, Thomas Pape (1996) relance l'ancien système de Böttcher (adopté aussi par Séguy) et élabore un système de phylogénétique chez les Sarcophagidae, sur la base de certaines analyses cladistiques. Il supprime la sous-famille Sarcophaginae sensu Rohdendorf & Auctoribus, en invoquant l'existence de nombreux groupes « monophylétiques » dans cette sous-famille. Ses opposants répondent que du point de vue taxonomique, en conformité avec les normes du Code International de Nomenclature Zoologique, chaque taxon, sans tenir compte de son rang, doit être « monophylétique », ou mieux dire « monotypique » et non « polyphylétique » ou « politypique ». Car, dans ce dernier cas, on ne peut garder un ordre systématique scientifique ou « naturel ».
Sous cette forme « phylogénétique », Pape a rétrogradé les genres de Sarcophagidae, établis dans les huit dernières décennies par les diptérologistes, en les considérant comme « sous-genres » ou « épithètes intercalées ».
Caractères morpho-taxonomiques des imagos
Pour la connaissance des Sarcophagidae certaisn étuduent donc leurs caractères morpho-taxonomiques. Ceux-ci peut donner non seulement la base de la compréhension des traits bio-écologiques générale de cette famille, mais aussi la possibilité de pénétrer dans la spécificité de ces éléments constitutifs.
Ces caractères permettraient également l’élaboration d’un instrument d’appréciation des limites de variabilité, qui coexistent dans les catégories taxonomiques fondamentales, dans une forme adéquate, scientifique et de niveau international.
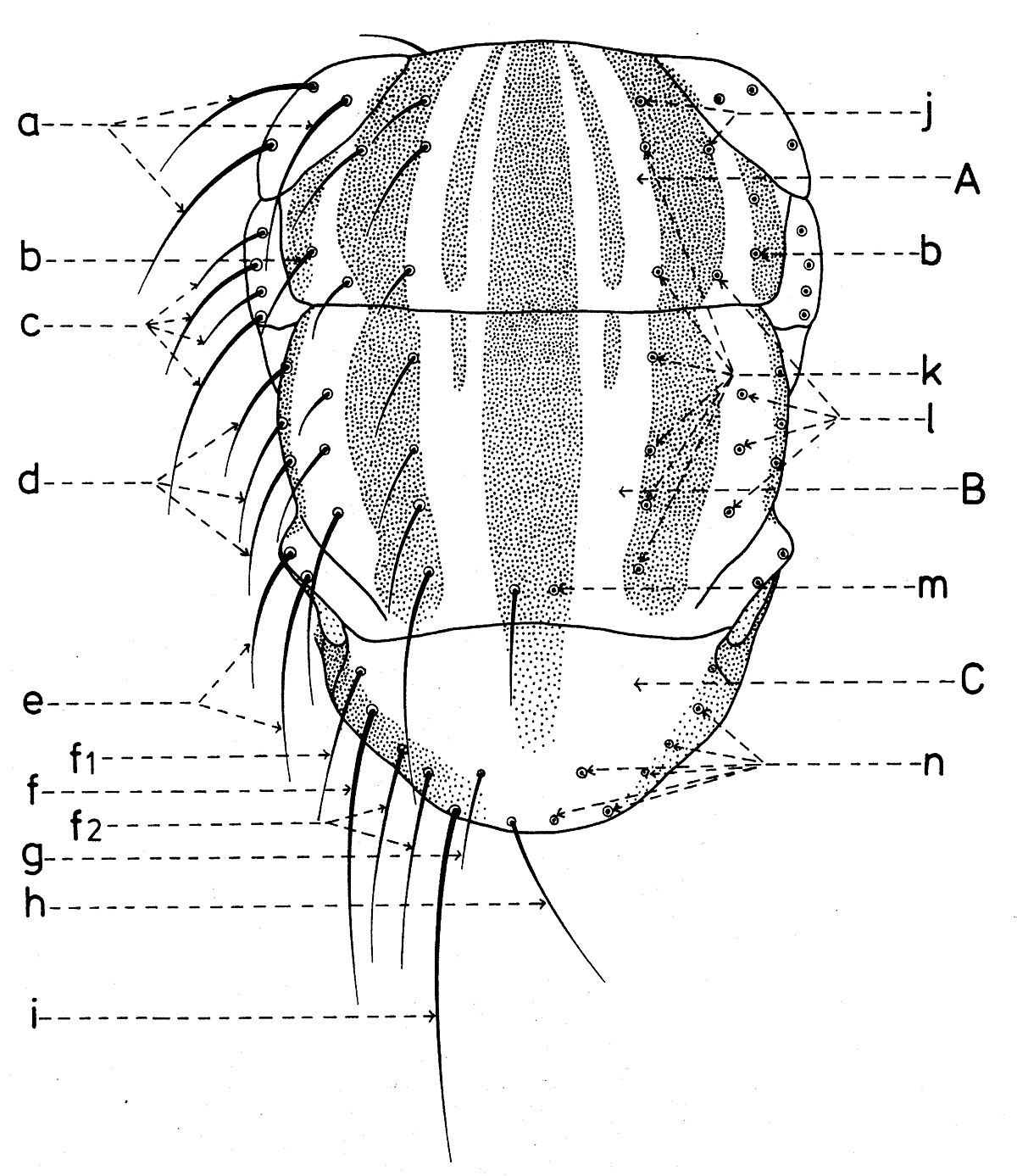
Chétotaxie de la tête
En général, les macrochètes portent les noms des parties dans lesquelles ils sont implantés. Ils forment, d’habitude, une garniture complète aux femelles (fig. 10). Chez les mâles (fig. 9) certains d’eux peuvent manquer (les macrochètes parafrontaux ou orbitaux), ou ils sont très peu développés (les macrochètes verticaux externes).
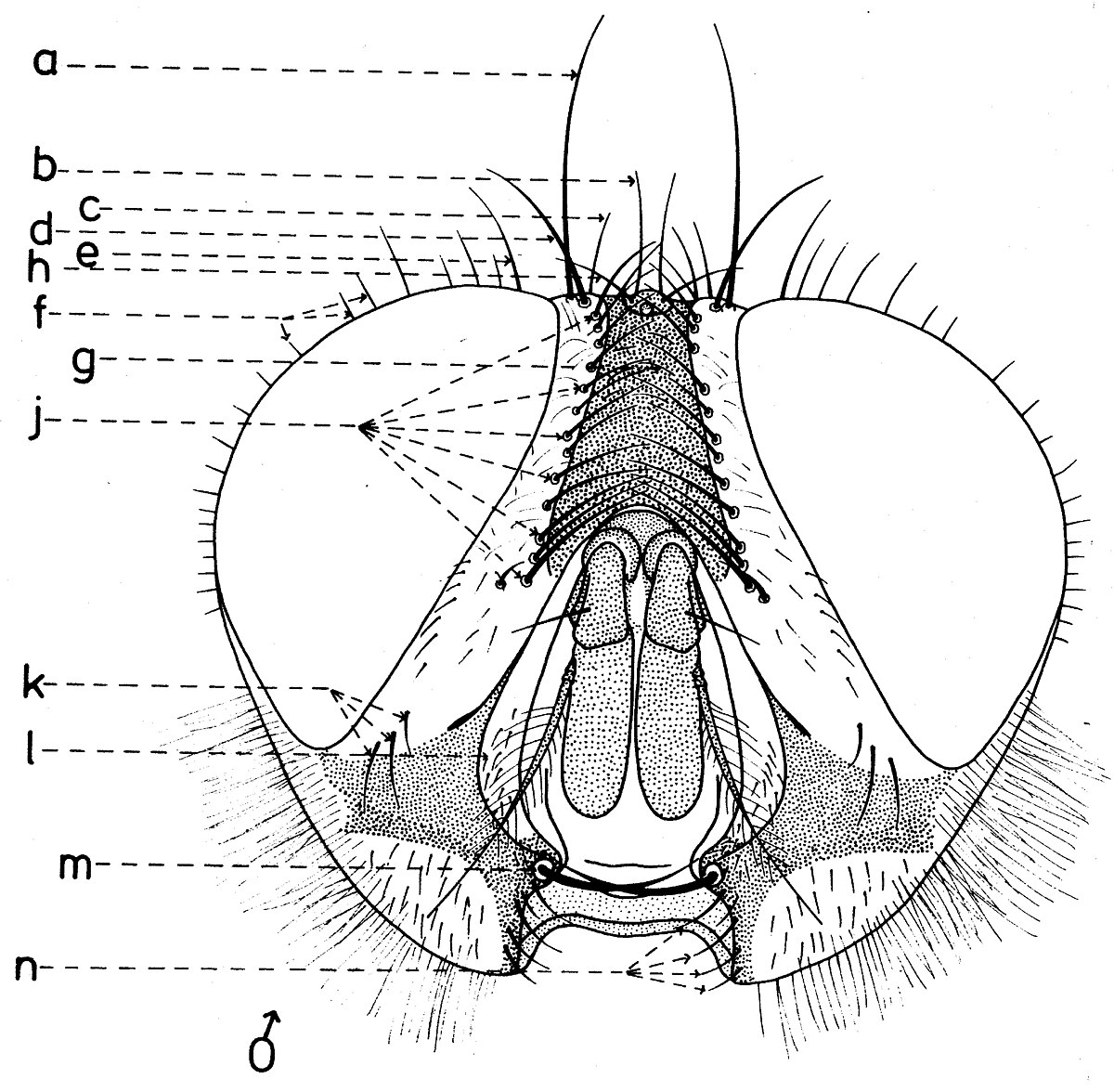
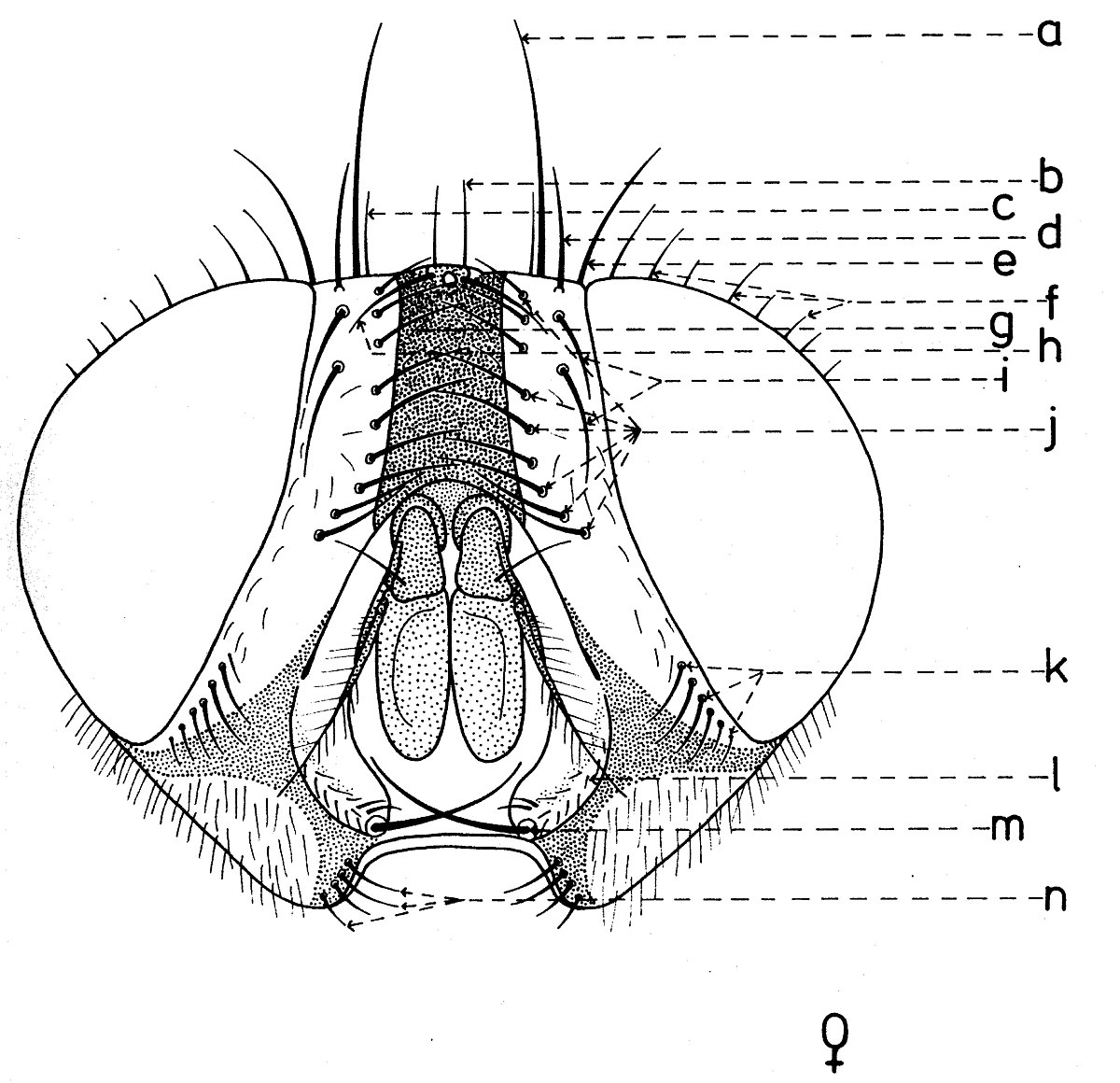
Sur le vertex, les plus forts sont les macrochètes verticaux internes (a), auprès des macrochètes verticaux externes (c), beaucoup plus petits ou qui manquent. Sur le triangle ocellaire, entre l’ocelle antérieur et ceux postérieurs sont les macrochètes ocellaires (h), plus petits, proclines et divergents, ayant parfois de dimensions très réduites. De chaque côté de la bande frontale médiane il y a 5-14 paires de macrochètes frontaux proclines (j), qui descendent du triangle ocellaire jusqu’au niveau du deuxième article de l’antenne, mais qui augmentent en grandeur et épaisseur. Même la bande frontale médiane a parfois quelques paires de macrochètes intrafrontaux (g), spécialement sur sa moitié postérieure.
Avant les macrochètes verticaux externes s’observe un macrochète prévertical (d) fort et rétrocline. Au femelles, sur les parafrontalies, à la proximité de la marge des yeux, il y a une paire de macrochètes parafrontaux (i) ou orbitaux bien développés et proclines. Sur les parafacialies et près de la marge antéro-inférieure des yeux on observe un rang vertical plus ou moins grand de macrochètes parafaciaux (k) ou gènes, dont ceux inférieurs sont souvent plus longs et plus gros que les autres. Au sommet de l’angle antéro-inférieure du péristome, connu sous le nom de vibrissarium, il existe un macrochète très développé qui s’appelle la grande vibrisse (m) et qui se croise avec sa symétrique avant la cavité buccale. Au-dessus d’elles, quelques petites vibrisses (l) montent plus ou moins sur les bordures faciales, jusqu’à un niveau caractéristique pour l’espèce. Sur les marges latérales de la cavité buccale se trouve un rang de macrochètes péristomaux (n), moins utilisés dans la taxonomie des Sarcophagides. Le péristome est couvert de poils noirs, mais parfois de poils blancs ou blancs jaunâtre aussi, qui descendent de la région postérieure de la tête.
Sur chaque côté de l’occiput (fig. ), à la hauteur des macrochètes verticaux (h, m) et ocellaires (f) se trouve un ou plusieurs macrochètes postverticaux (l) et postocellaires (k) piliformes. À la marge postérieure de la bordure postoculaire il y a un rang de macrochètes postoculaires (i) proclines de dimensions réduites et, plus bas, 2-3 rangs de microchètes occipitaux (j), disposés sur la moitié supérieure de la région postérieure.
Biométrie de la tête
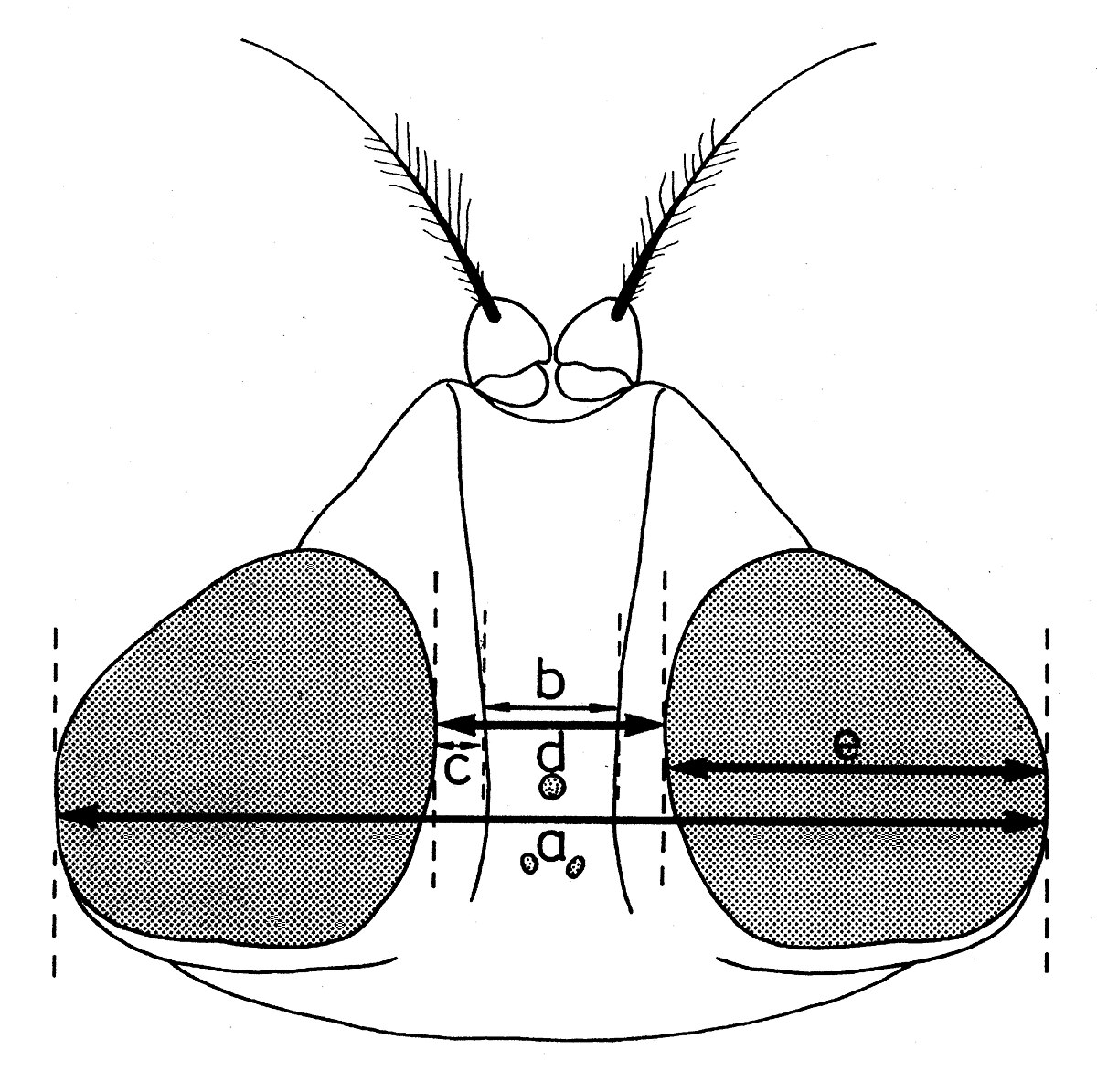
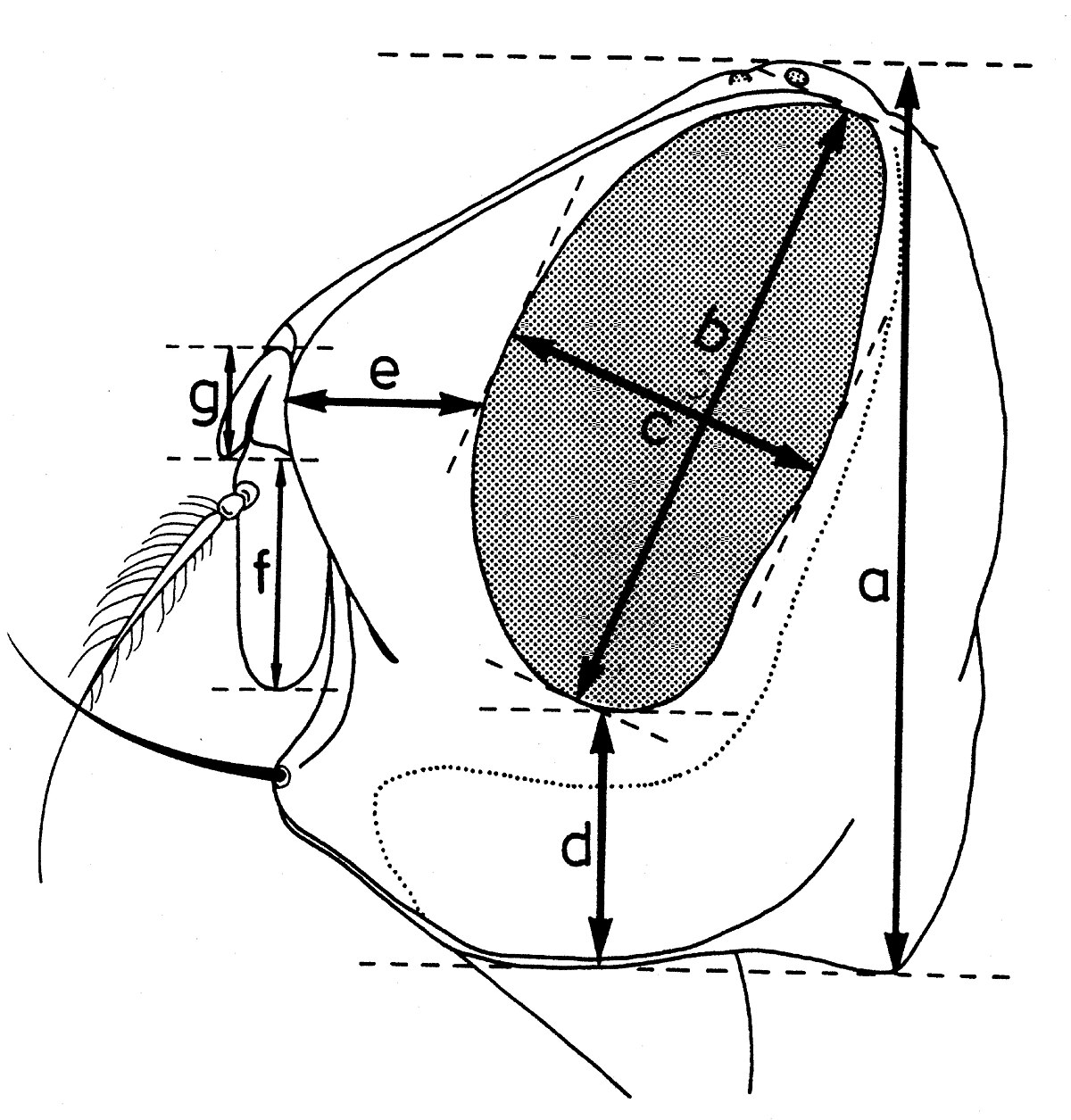
Dans la taxonomie des Sarcophagides, comme dans celle d’un grand nombre de diptères, certains indices biométriques ont une importance assez grande. Les facteurs suivants sont pris en considération :
a - Les indices établis quand la tête est examinée du dessus (fig. 11):
- largeur de la tête (a);
- largeur d’un œil (e);
- largeur du front (d), mesurée au lieu le plus étroit;
- largeur de la bande frontale médiane (b), mesurée au même niveau;
- largeur d’une parafrontalie (c), mesurée au même lieu.
b - Indices déterminés quand la tête est examinée de profil (fig. 12):
- hauteur de la tête (a);
- grand diamètre oculaire (b);
- petit diamètre oculaire (c);
- hauteur du péristome (d);
- proéminence frontale ou profrons (e);
- rapport entre la longueur du troisième article de l’antenne (f) et celle du deuxième article (g).
Chétotaxie du thorax
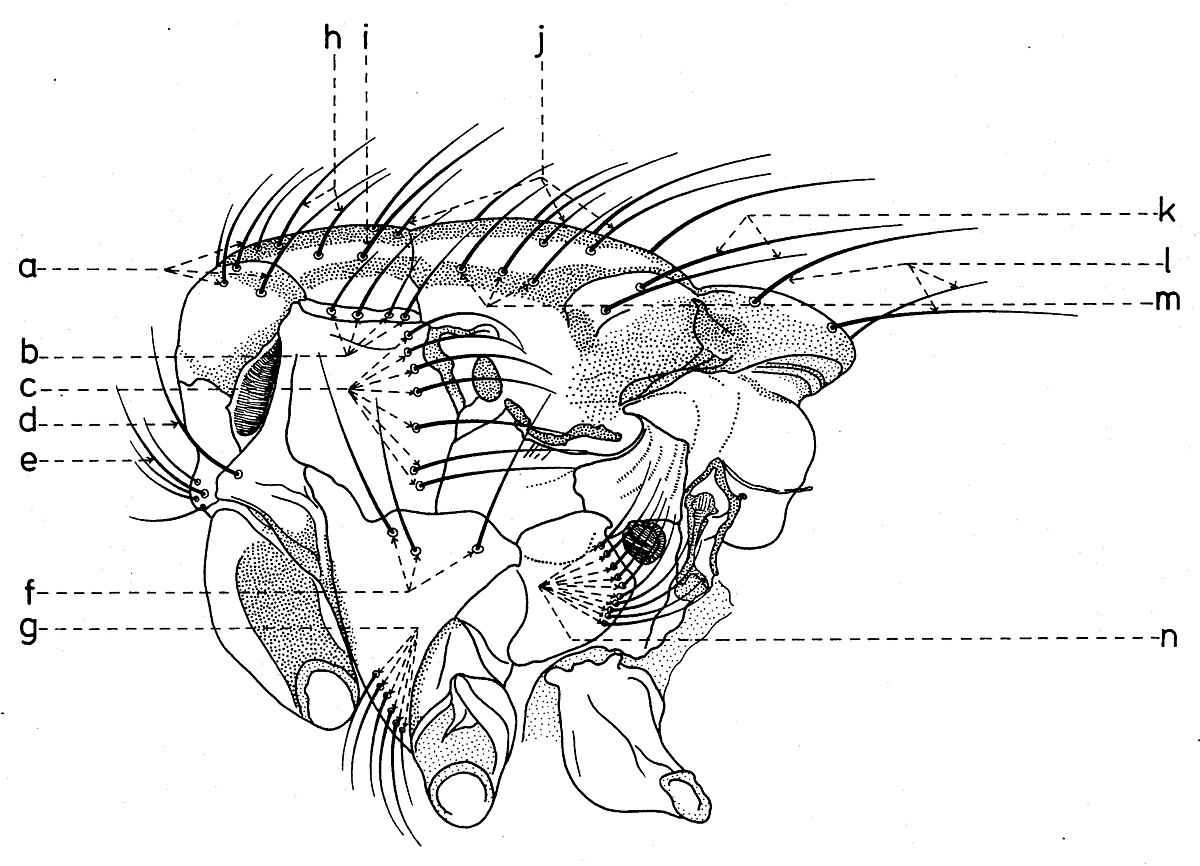
Avec peu d’exceptions, les macrochètes du thorax portent aussi les noms des lieux ou ils sont fixés. Sur sa partie dorsale (fig. 13) s’observent quelques rangs et groupes de:
- macrochètes achrosticales (m). Souvent ils peuvent être limités seulement à une paire préscutelaire; sur la partie antérieure du mesonotum peuvent exister encore quelques paires courtes, qui dépassent un peu la pilosité fondamentale du tegumentum.
- macrochètes dorso-centraux (k) présuturaux au nombre de 3-4 et postsuturaux au nombre de 3-5 paires. Les derniers ont une grande importance dans la taxonomie du groupe.
- macrochètes intraalaires (l). Celui présutural (b) peut manquer, et les autres sont au nombre variable.
- macrochètes huméraux (a) sur le callus humeral.
- macrochètes posthumeraux (j).
- macrochète présutural (b). À la famille Sarcophagidae, de règle, il se trouve plus bas et plus vers l’extérieur devant le dernier macrochète posthumeral. Parfois il peut manquer.
- macrochètes supraalaire (d), situés après la suture et au-dessus de l’aile.
- macrochètes notopleuraux (c) sur les notopleures.
- macrochètes postalaires (e) sur les callus postalar.
- macrochètes scutellaires (n) qui se divisent en: macrochètes apicaux (h) - présents ou absents, petits, demi-érects, croisés, divergents etc.; macrochètes subapicaux (i) - très développés; les macrochètes latéraux - qui se catégorisent en: macrochètes latéraux proprement dit (f2) - au nombre de 1-3 ou peuvent manquer, macrochètes basaux (f) - forts et macrochètes prébasaux (f1); macrochètes discaux (g) - placés au-dessus du disque scutellaire et près de l’apex, plus fins et d’habitude demi-érects.
De profil (fig. 14), le thorax présent :
- macrochètes propleuraux (e), souvent entourés de quelques poils macrochètiformes supplémentaires.
- macrochètes prostigmaticaux (d), entourés aussi de 1-2 poils auxiliaires.
- macrochètes mésopleuraux (c).
- macrochètes sternopleuraux (f), situés souvent en ligne, dans la position 1:1:1; mais le médian peut manquer et alors leur position est 2:1 ou 3:1 (Agria Robineau-Desvoidy, Angiometopa Brauer & Bergenstamm).
- macrochètes infrasternopleuraux (g).
- macrochètes hypopleuraux (n).
Nervation
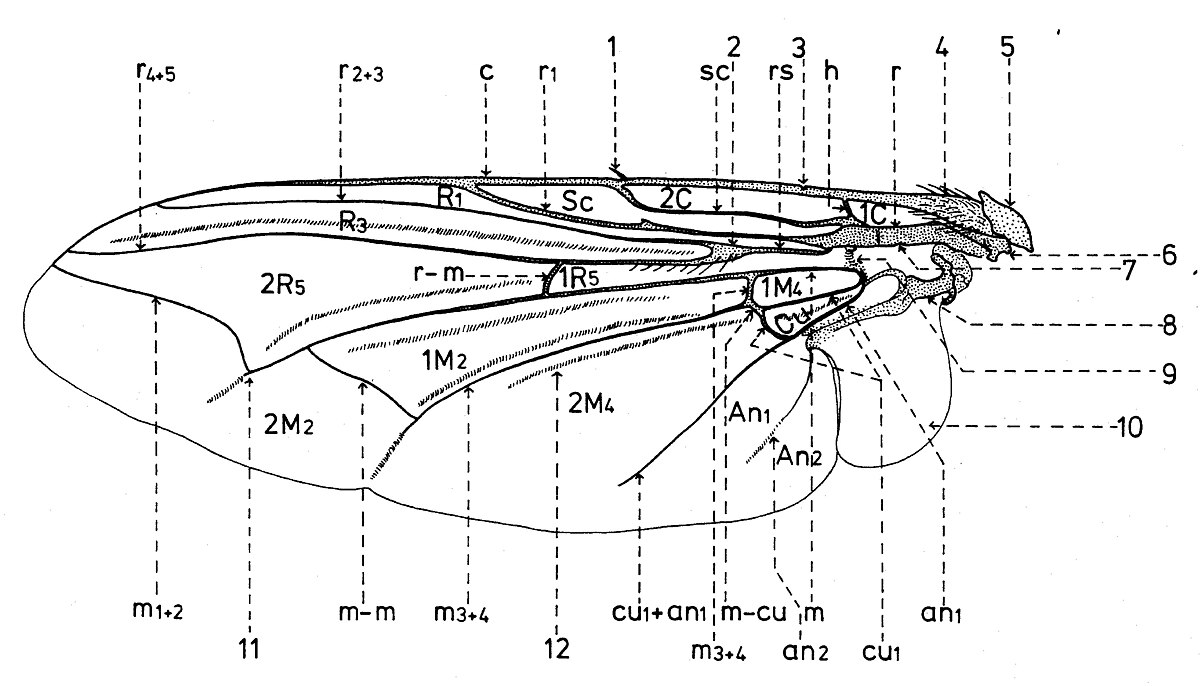
La disposition des nervures (longitudinales et transversales ou nervules) est semblable avec celle des autres familles des superfamilles Sarcophagidea et Tachinidea (fig. 15).
D’après le système de Comstock-Needham, les nervures longitudinales qui se trouvent chez les Sarcophagides, sont les suivantes:
- la nervure costale (c), qui confine la partie antérieure de l’aile jusqu’à l’apex. Elle a deux fractures dans sa partie proximale; à une d’elle se trouve l’épine costale sensorielle ou plectrum (l). Elle se dilate à la base, portion nommée costagium (4); et au point d’insertion de la nervure costale il y a deux sclérites sous la forme d’écailles: un proximal ou épaulette qui est une tegula et l’autre distal ou basicosta (5). Dans sa monographie sur les Sarcophagides paléarctiques, Verves fait toujours la confusion entre ces deux sclérites, en les inversant.
- la nervure subcostale (sc) est courte, concave et atteinte la nervure costale au niveau de l’épine costale.
Le système des nervures radiales est l’élément le plus fort de l’aile et se divise en :
- la nervure r1 ou la première nervure radiale (r1), convexe et prolongée jusqu’à l’extrémité de l’aile. Parfois elle est pourvue de macrochètes à la base ;
- la nervure r2+3 ou la deuxième et la troisième nervure radiale (r2+3) ;
- la nervure r4+5 ou la quatrième et la cinquième nervure radiale (r4+5), souvent pourvus de microchètes du nœud radial (2) et sur une distance plus longue ou plus courte vers la nervure transversale r-m (r-m).
Le système des nervures médiales (m), en connexion avec celui des nervures cubitales, s’unis aussi à la base avec les nervures radiales, par l’intermédiaire d’une nervure moins distincte, nommée arculus (9). Ainsi, elles sont dépendantes tant du tronc radial commun ou brachiolum (7), que du celui du quatrième sclérite axillaire (8). Elles se pressentent comme :
- la nervure médiale (m), qui se dirige vers l’extrémité de l’aile, où se courbe en haut et, puis, se courbe vers la marge antéro-apicale sous la forme de la nervure m1+2 ou la première et la deuxième nervure médiale (m1+2). Le point de courbure angulaire se nom cubitulus (11), celui-ci étant prolongé soit d’un pli, soit d’une nervure très courte nommée appendix.
- la nervure m3+4 ou la troisième et la quatrième nervure médiale (m3+4), qui se bifurque transversalement près de la base et, puis, se dirige vers la marge postérieure de l’aile, vers l’excision préanale.
Viennent aussi :
- la nervure cu1+an1 ou la première nervure cubitale et la première nervure anal (cu1+an1), qui marchent indépendamment sur une petite portion basale (cu1 et an1) et puis s’unissent.
- la nervure an2 ou la deuxième nervure anale (an2).
Les nervures transversales sont au nombre de 4, à savoir,
- la nervure transversale humérale (h), qui unie près de la base les nervures costale et subcostale.
- la nervure transversale radio-médiale (r-m), qui unie les nervures et m.
- la nervure transversale intermediale (m-m), qui unie les nervures m et m3+4.
- - la nervure transversale médio-cubitale (m-cu), qui unie à la base les nervures m3+4 et cu1.
Les cellules sont les espaces alaires délimitées par nervures et portent d’habitude les noms des nervures qui les confinent dans leur partie antérieure. Elles sont notées avec de lettres majeures et peuvent être fermées (quand elles sont délimitées apicalement par de nervures transversales ou l’espace cellulaire est entouré par deux nervures qui confluent. En cas contraire, elles sont ouvertes.
Chez les Sarcophagidae sont présentes les cellules suivantes:
- la cellule costale basale (1C);
- la cellule costale distale (2C);
- la cellule subcostale (Sc);
- la première cellule radiale ();
- la troisième cellule radiale (R3);
- la cinquième cellule radiale basale (1R5);
- la cinquième cellule radiale distale (2R5);
- la deuxième cellule médiale basale (1M2);
- la deuxième cellule médiale distale (2M2);
- la quatrième cellule médiale basale (1M4);
- la quatrième cellule médiale distale (2M4);
- la cellule cubitale (Cu);
- la première cellule anale (An1);
- la deuxième cellule anale (An2).
Abdomen des Sarcophagidae
Hormis les caractères chétotaxiques et chromatiques des Sarcophagidés, les plus importantes caractères taxonomiques sont ceux des structures morphologiques de l’abdomen et notamment du postabdomen. Pour les comprendre ont doit les examiner, sous la prisme des recherches modernes.
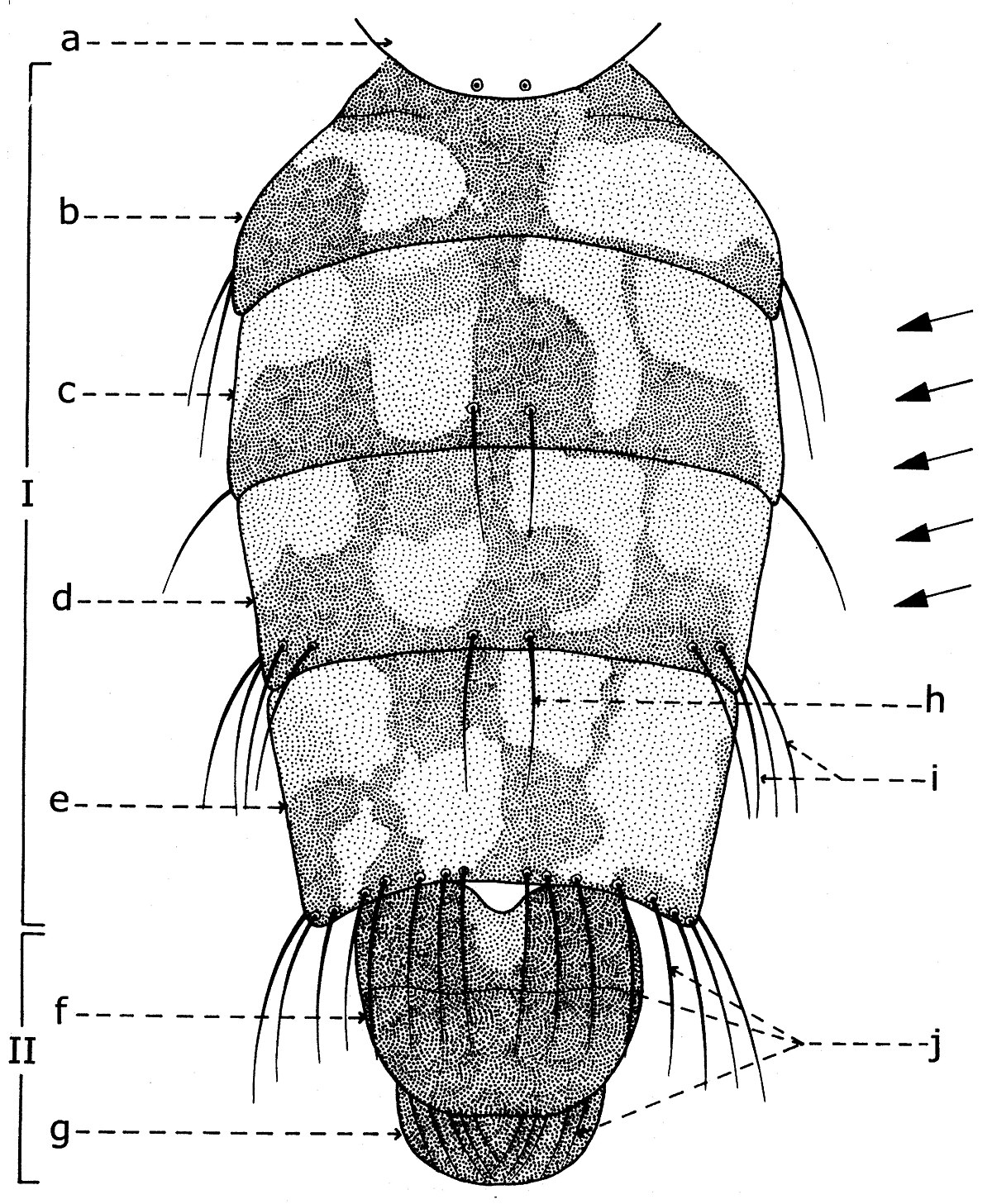
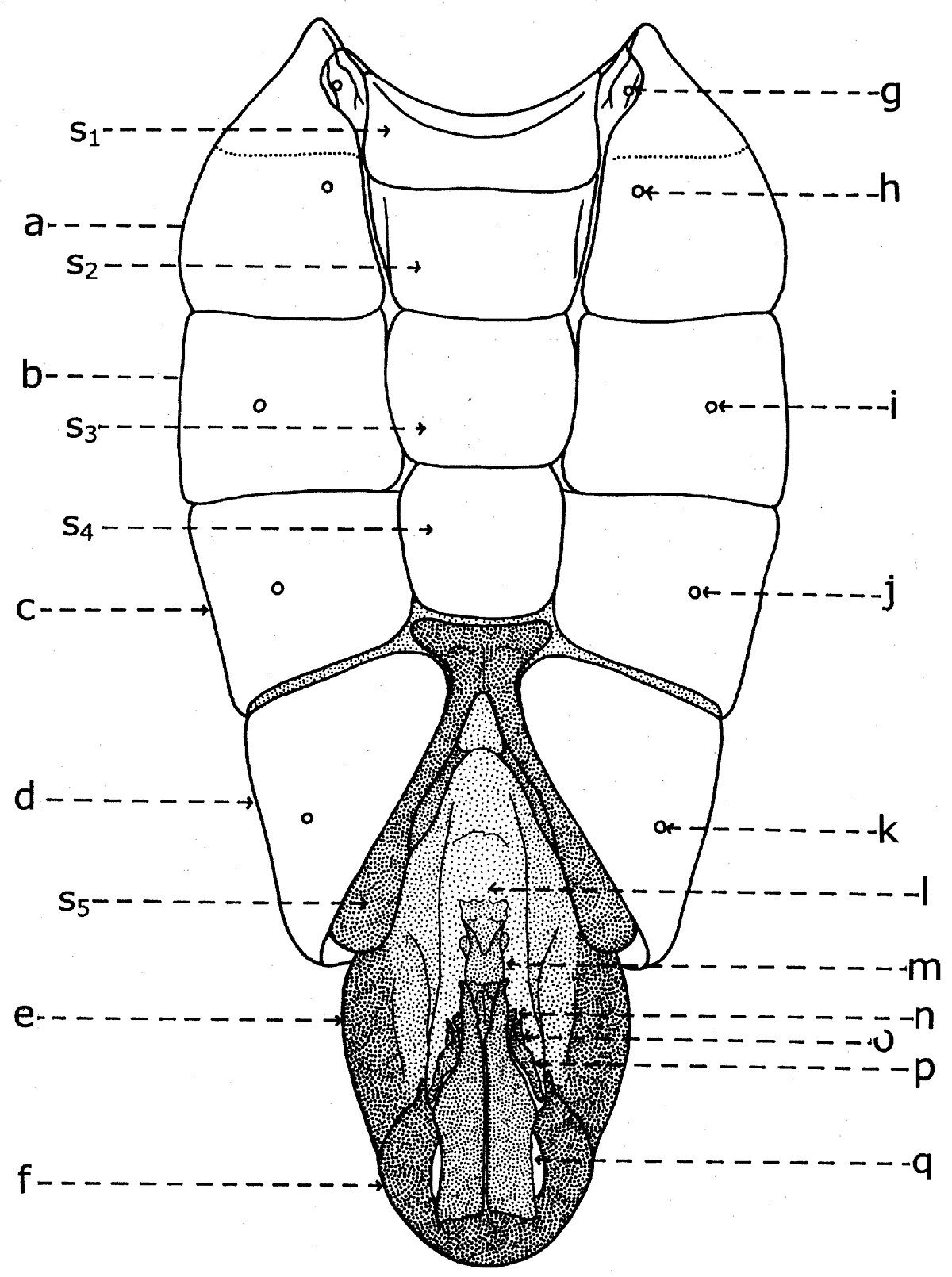
Théoriquement, l'abdomen des Sarcophaginae est formé de 11 segments, dont 5 segments abdominaux visibles constituent le préabdomen (fig. 16, I; fig. 17, b-d) et les 6 segments réduits, atrophiés et transformes en aide des fonctions de reproduction, représentent le postabdomen (fig. 16, II; fig. 17, f-g).
Le préabdomène
Le préabdomen (fig. 16, I) est allongé-ovoidal ou svelte chez le mâle et plus large chez la femelle. Ses dessins dorsaux sont, le plus souvent, formés de taches changeantes de tomentum argenté, qui alternent avec de taches noires, donnant l'aspect en damier. Ces dessins, caractéristiques à la majorité des espèces de la sous-famille, sont variables, d'après l'incidence de la lumière. Mais, de plusieurs fois, ces taches sont disposées en bandes dorsales longitudinales sur tous les tergites préabdominaux. Autrefois, le préabdomen est cendré, avec taches plus ou moins circulaires et allongées sur tous les tergites ou seulement sur une partie de ceux-ci (l'espèce paléarctique Yerohama maculata Meigen, ou le genre éthiopienne Poecilometopa Villeneuve). Parfois, les bandes noires dorsales sont disposées transversalement et, très rarement, le préabdomen est complètement noir.
Aux mâles, les sternites III et IV (fig. 17, s3-s4) sont plus ou moins semblables au sternite II, bien qu'ils donnent l'impression qu'ils sont plus petits et plus étroits, tandis que chez les femelles, les sternites III-IV sont approximativement de la même forme. Le sternite V des mâles (fig. 10, s5) est hétéromorphe, ayant une pièce basale et deux lames latérales, qui peuvent former une fenêtre, souvent triangulaire et nommée fenestella. Chez beaucoup d'espèces, mais surtout chez celles néotropicales, il a des prolongements digitiformes ou d'autres apophyses simples et bifides. Sur la marge antérieure des lames latérales, comme sur leur surface inférieure, se trouvent un grand nombre de poils assez longs et, même, très longs vers leur apex. Beaucoup d'espèces ont de grandes agglomérations de macrochètes récurrents, courts et fournis sur les marges lamellaires intérieures, nommées brosses.
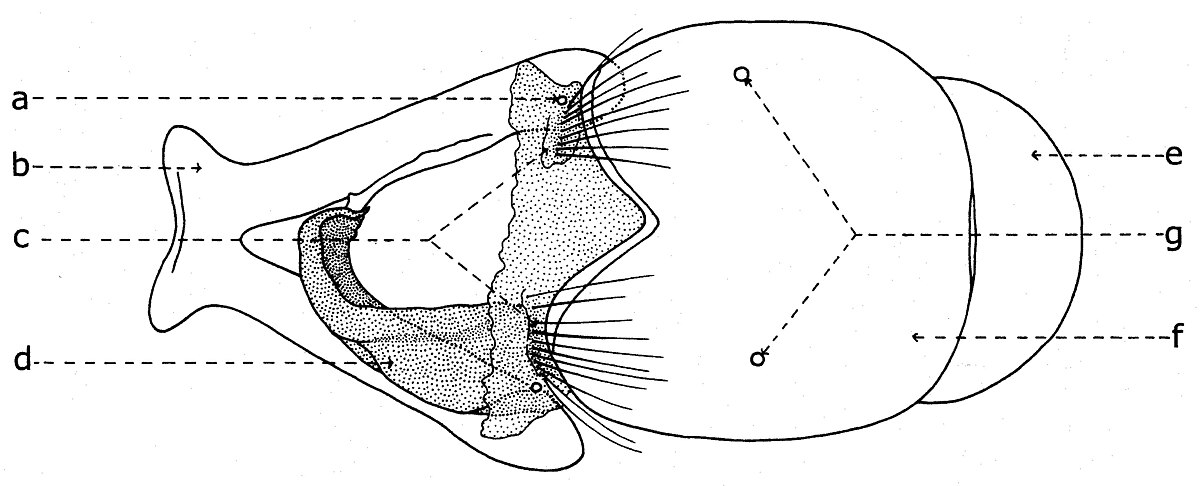
Le postabdomen
Le postabdomen du mâle est connu sous le nom de terminalia mâle ou hypopigium (fig. 16, II; fig. 17, e-f; fig. 18, e-f; fig. 19, e-f). Après l'éclosion des imagos, le hypopigium soufre une rotation de 360 degrés par rapport à l'axe longitudinal de l'abdomen, entre les segments V et VI. Ainsi le canal éjaculateur se tourne autour de l'intestine postérieure, le postabdomen revenant apparemment dans sa position initiale normale. Ce type de hypopigium circumversum entraîne après lui diverses modifications et asymétries des certaines sclérites (par exemple, le sternite VI), la fusion ou la disparition des autres, avec conséquences profondes dans sa morphologie générale (la migration des segments et spiracles etc.).
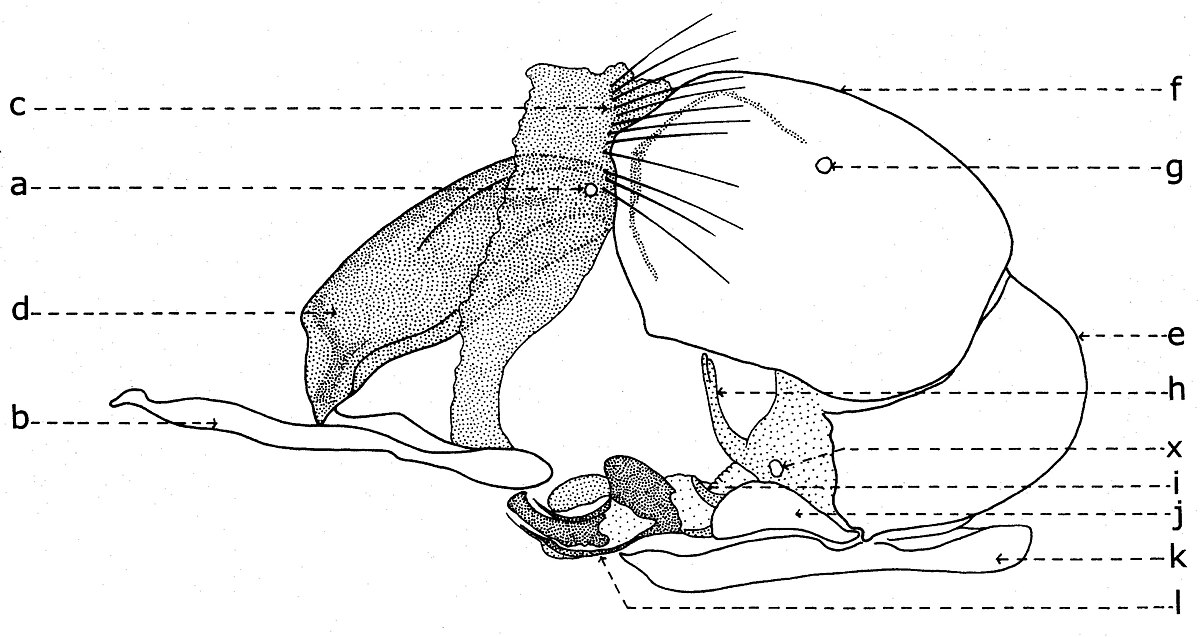
Chez beaucoup de représentants de la sous-famille Sarcophaginae, le segment VI abdominal ou le premier segment postabdominal (fig. 18-19) a un tergite entier ou divisé en deux hemitergites (c) réduits et pas sclérifiés, identifiables (dans la plupart des cas) par deux groupes linéaires de macrochètes. Le sternite VI (d) est déformé, asymétrique, ayant développée seulement la partie gauche et il est soudé de la base des lobes latérales du sternite V (b). Dans la membrane pleurale sont visibles les spiracles de la paire VI (a). Cependant, aux autres genres et espèces, le tergite VI peut manquer complètement (Bercaea cruentata Meigen, Blaesoxipha krameri Baranov).
Après le tergite VI se succède un tergite bien développé, qui est considéré le syntergite VII+VIII (fig. 19, f), parce qu'il résulte de la fusion des tergites VII et VIII. La preuve est donné tant par l'existence d'une suture sur son milieu, que par la présence des spiracles de la paire VII (g) seulement dans la région correspondante au tergite VII, l'autre paire (la VIIIme) ne se trouve pas chez les imagos des diptères. Il est nommé aussi le tergite génital ou le premier tergite génital.
A une série d'espèces de Sarcophagides, le tergite VI est soudé avec le tergite VII+VIII, sa ligne de suture étant marquée visiblement (Agria affinis Fallen, Wohlfahrtia balassogloi Portschinsky). En échange, à une autre série, les tergites VI, VII et VIII sont plus ou moins individualisés (Mimodexia magnifica Rohdendorf).
Se succède le tergite du dernier segment, connu sous le nom de tergite anal (e), le deuxième tergite génital ou, le plus souvent, epandrium. Zumpt et Heinz (1949,1950) l'ont conçu comme « 10th segment and cerci ». Il est le tergite auquel se connecte, dans sa partie antéro-ventrale, l'organe copulateur mâle, avec tous les sclérites que lui desservent intimement et, dans la partie postérieure, ses appendices auxiliaires.
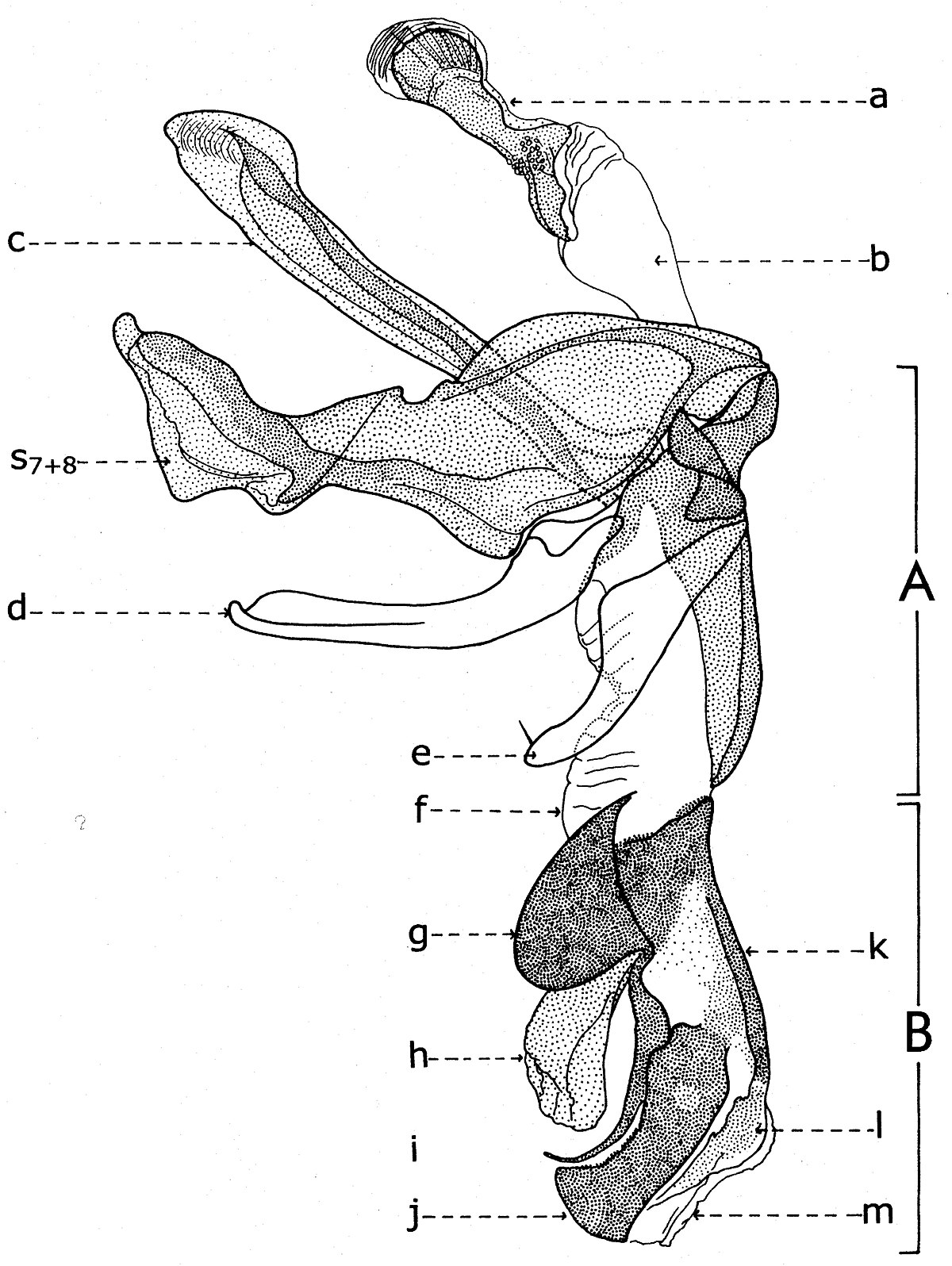
Explication de la fig. 20. Phallosome de Sarcophaga carnaria Bottcher., vu de profil. A: basiphallus; B: distiphallus; a: apodeme du canal ejaculateur; b: canal ejaculateur; c: phallapodema; d: pregonite; e: postgonite; f: membrana; g: lobes hypophalliques basaux; h: lobes membranaux; i: lobes hypophalliques ventraux; j: styles; k: paraphallus; l: branches laterales du paraphallus; m: juxta; S7+8: sternite VII+VIII.
A la base de l'organe copulateur se trouve un sclerite relativement grand (fig. 20, st7+8), qui l'entoure et a un aspect d'un auget. Il est connu dans littérature sous le nom de tergosternum IX ou hypandrium et défini, en général, comme le sternite IX (Tuxen, 1970). Mais Zumpt et Heinz croient que dans sa structure participent tant le tergite IX, que le sternite IX, d'où la dénomination prise de Patton et Cushing. Séguy (1941) n'a donné aucune attention à cette formation, indiquant erronément le sternite IX sous le nom de l'apodème du canal ejaculateur (a).
Dans une interprétation actuelle, il n'est ni le « sternite IX » et ni le résultat de la fusion des tergite et sternite IX. Il est simplement le correspondent du tergite VII+VIII, ou mieux dire, le sclerite qui résulte de la fusion des sternites VII et VIII.
Tenant compte du postulat de Weber, d'après lequel l'orifice génital se trouve au bout postérieur du sternite IX, alors on peut constater que l'orifice génital n'est pas au bout du « tergosternum IX » ou hypandrium (sensu Emden et Hennig) - comme pensent encore les dipterologiste contemporains, mais au celui d'une apophyse sous la forme d'une baguette aplatisse, située dans l'intérieur du sternite VII+VIII et appréciée erronément par les auteurs comme une néoformation, la phallapodema ou l'apodème du pénis (c). Dans autres termes, comme s'est observé autrefois aussi aux Syrphidae (Lehrer, 1971), cette phallapodème est le véritable sternite IX.
D'un côté et d'un autre de l'organe copulateur (fig. 20) il y a une paire d'appendices plus ou moins sous la forme de crochètes : les prégonites ou les paramères antérieurs (d) et les postgonites ou les paramères postérieurs (e). Leur origine n'est pas bien clarifiée par les auteurs, la majorité acceptant qu'ils dérivent du sternite IX (sensu auct.). Zumpt et Heinz affirment, sans aucune preuve, qu'alors quand il existe deux paires de paramères (comme dans le cas des Sarcophagidae), ils proviennent de la division des lobes paramèraux. Mais, des recherches actuelles sur le postabdomen des Syrphidae résulté que les prégonites ont leur origine dans le sternite VIII (sternite IX sensu Emden & Hennig), tandis que les postgonites proviennent du sternite IX (phallapodema sensu Emden & Hennig).
Dans la partie postérieure du tergite X sont fixées deux paires d'appendices relativement longs: les paralobes (fig. 19, j) et les cerques (fig. 19,k). La première paire, désignées par divers auteurs sous le nom de mesocerques latéraux, forceps externes, gonopodes ou surstyli, descendent du sternite X et pas du tergite du segment IX (conf. Emden & Hennig et. Auct.). Chez toutes les espèces étudiées de la famille Syrphidae, les branches ou les prolongements postérieurs du sternite X sont soudes toujours avec ces paralobes.
Chez Sarcophaga carnaria Boettcher., le sternite X est très réduit, en se présentant comme des hemisternites (fig. 19, x), plus ou moins ovoïdaux, petits et situes dans la membrane d'entre les paralobes et la marge inférieure du tergite X. Pour être découvertes, il est nécessaire une forte détente de la membrane ou le relèvement des paralobes qui les couvrent. Le sternite X a été observé tout d'abord par Patton (1934), qui l'a considéré erronément comme le spiracle de la VIII-me paire et, puis, par Zumpt et Heinz, qui ont lui donné l'interprétation correcte.
En échange, les cerques sont les réminiscences du segment XI abdominal, duquel n'est pas resté que le proctiger ou la membrane qui s'étend dans l'émargination du tergite X, y compris seulement l'anus.
Les genres des Sarcophagidae selon les morphologistes
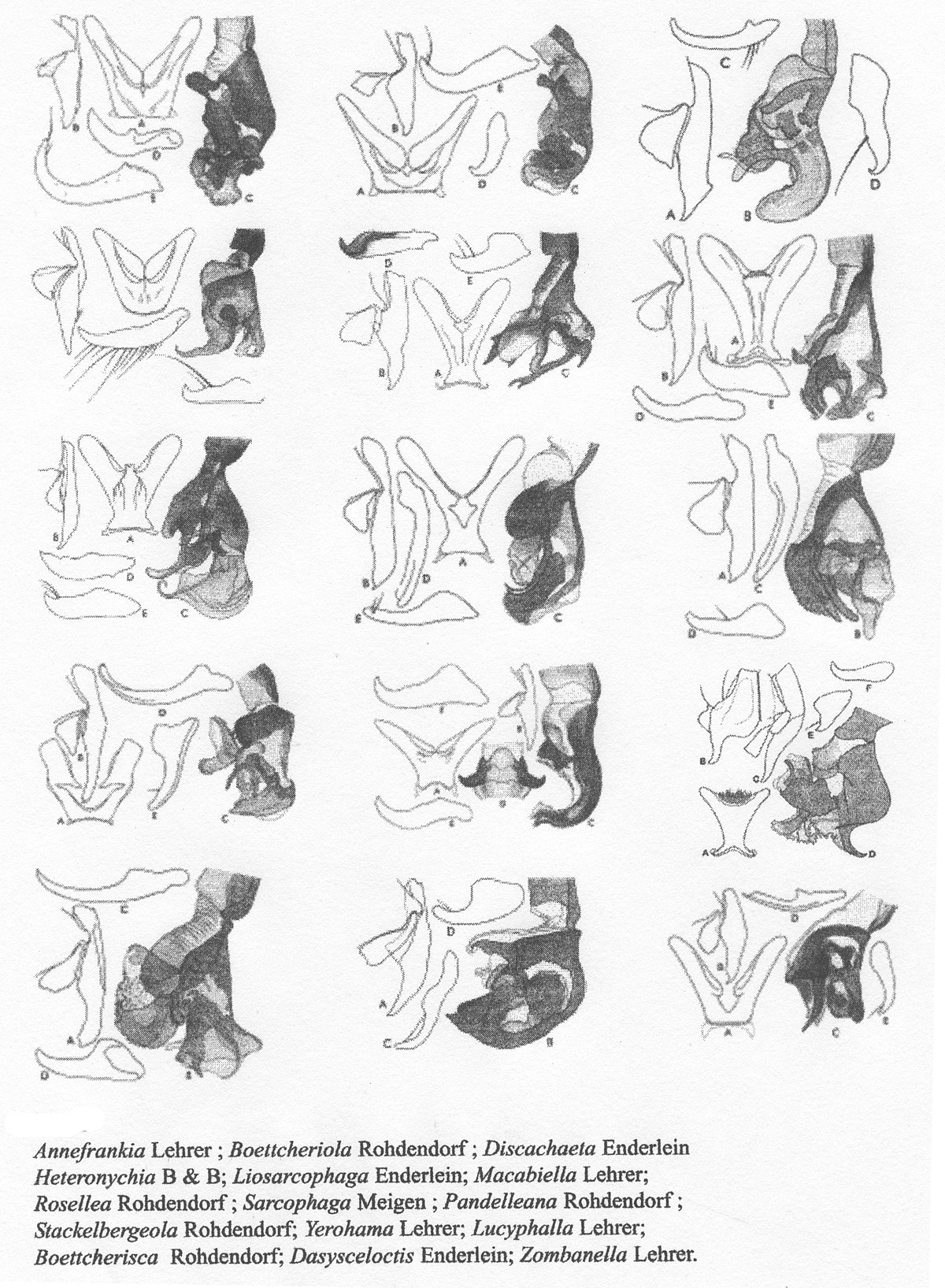
Les genres de cette famille sont très nombreux dans toutes les régions zoogéographiques. En même temps, ils ne sont pas assez bien connus et nous ne savons pas dans quelle proportion ils représentent une certaine région. C’est pourquoi, nous donnons une liste avec les genres les mieux représentés dans les régions paléarctique, afrotropicale et orientale.
- Aethianella Zumpt
- Aethiopisca Rohdendorf
- Afrohelicobia Zumpt
- Afrothyrsocnema Rohdendorf
- Agria Robineau-Desvoidy
- Agriella Villeneuve
- Aleximyia Rohdendorf
- Amharomyia Verves
- Amibia Robinea-Desvoidy
- Anacanthothecum Rohdendorf
- Angiometopa Brauer & Bergenstamm
- Annefrankia Lehrer
- Anthostilophalla Lehrer
- Apodacra Macquart
- Asceloctella Enderlein
- Asiometopia Rohdendorf
- Aspidophalla Rohdendorf
- Bantuphalla Lehrer
- Baranovisca Lopes
- Barnardia Lehrer
- Batissophalla Rohdendorf
- Bechuanella Lehrer
- Bercaea Robinea-Desvoidy
- Beziella Enderlein
- Bilenemyia Verves
- Blaesoxipha Loew
- Boettcherella Enderlein
- Boettcherisca Rohdendorf
- Brachicoma Rondani
- Brasia Strand
- Burmanomyia Fan
- Callostuckenbergia Lehrer & Lehrer
- Camerounisca Verves
- Ceratophalla Rohdendorf
- Cercosarcophaga Zumpt
- Chaetanicia Townsend
- Chaetophalla Rohdendorf
- Craticulina Bezzi
- Ctenodasypygia Enderlein
- Ctenodasypygia Enderlein
- Curranea Rohdendorf
- Curranisca Rohdendorf
- Curtophalla Lehrer
- Cyclophalla Rohdendorf
- Cylindrothecum Rohdendorf
- Danbeckia Lehrer
- Dasysceloctis Enderlein
- Deconinckia Lehrer
- Devriesia Lehrer
- Dinemomyia Chen
- Diplonophalla Lehrer
- Dolichotachina Villeneuve
- Dovporiella Lehrer
- Draculana Lehrer
- Durbanella Lehrer
- Engelisca Rohdendorf
- Eremasiomyia Rohdendorf
- Eumetopiella Hendel
- Fanzidella Lehrer
- Fengia Rohdendorf
- Golania Lehrer
- Goniophyto Townsend
- Grimaldiomyia Lehrer
- Hadashophalla Lehrer
- Harpagophalla Rohdendorf
- Harpagophalloides Rohdendorf
- Helicophagella Enderlein
- Heteronychia Brauer & Bergenstamm
- Hilarella Rondani
- Hoa Rohdendorf
- Hoplacephala Macquart
- Hoplocephalomima Verves
- Horia Kano, Field & Shinonaga
- Horisca Rohdendorf
- Hyperacanthisca Rohdendorf
- Ihosyia Verves
- Imerina Lehrer
- Iranihindia Rohdendorf
- Jaennickeana Lehrer
- Jantiella Rohdendorf
- Kanomyia Shinonaga & Tumrasvin
- Kramerea Rohdendorf
- Krombeinomyia Verves
- Leclercqiomyia Lehrer
- Leucomyia Brauer & Bergenstamm
- Liopygia Enderlein
- Liosarcophaga Enderlein
- Livingstonelia Lehrer
- Macabiella Lehrer
- Macronychia Rondani
- Malawithyyrsia Lehrer
- Malliophalla Lehrer
- Mandalania Lehrer
- Mauritiella Verves
- Mediterranisca Rohdendorf
- Mesomelaena Rondani
- Metopia Meigen
- Metopodia Brauer & Bergenstamm
- Miltogramma Meigen
- Miltogrammidium Rohdendorf
- Miltogrammoides Rohdendorf
- Mitumbana Lehrer
- Nepalisca Rohdendorf
- Nguyenia Lehrer
- Nuzzaciella Lehrer
- Nyctia Robineau-Desvoidy
- Nyikamyia Lehrer
- Oebalia Robineau-Desvoidy
- Pandelleana Rohdendorf
- Pandelleisca Rohdendorf
- Parabellieria Verves
- Parasarcophaga Johnston & Tiegs
- Petuniophalla Lehrer
- Phallocheira Rohdendorf
- Phallosphaera Rohdendorf
- Pharaonops Lehrer
- Phrosinella Robineau-Desvoidy
- Phylloteles Loew
- Pierretia Robineau-Desvoidy
- Prionophalla Rohdendorf
- Protomiltogramma Townsend
- Pterella Robineau-Desvoidy
- Pterolobomyia Lehrer
- Radamaia Lehrer
- Ravinia Robineau-Desvoidy
- Robertiana Lehrer
- Robineauella Enderlein
- Rohdendorfisca Grunin
- Roselles Rohdendorf
- Salemiophalla Lehrer
- Sarcophaga Meigen
- Sarcophila Rondani
- Sarcorohdendorfia Baranov
- Sarcosolomonia Baranov
- Sarcotachina Portschinsky
- Sarcotachinella Townsend
- Seniorwhitea Rohdendorf
- Senotainia Macquart
- Servaisia Robineau-Desvoidy
- Seselwana Lehrer
- Sinonipponia Rohdendorf
- Spatulapica Fan
- Sphenometopia Townsend
- Stuckenbergiella Verves
- Synorbitomyia Townsend
- Taxigramma Peris
- Thomsonea Rohdendorf
- Transvaalomyia Lehrer & Lehrer
- Tuberomembrana Fan
- Virungana Lehrer
- Wittemyia Lehrer
- Wohlfahrtia Brauer & Bergenstamm
- Wohlfahrtiodes Villeneuve
- Xanthopterisca Rohdendorf
- Yerohama Lehrer
- Zombanella Lehrer
- Zumptiopsis Lehrer & Lehrer
- Zumptisca Rohdendorf
Sources bibliographiques principales
- Baranov, N. (1925), Neue Dipteren aus Serbien. – Let. Pol. Odl. Kontr. Sta. Topcideru, Belgrad, 1:1-11.
- Baranov, N. (1941), Zweiter Beitrag zur Kenntnis der Gattung Sarcophaga (s.l.). Vet. Arh., 11:361-404 [In Croatian and German].
- Becker, T. (1908), Dipteren der Kanarischen Inseln. Mitt. zool. Mus. Berlin, 4:1-180.
- Böttcher, G. (1912), Die männlichen Begattungswerkzeuge der Arten bei dem Genus Sarcophaga Meigen. und ihre Bedeutung für die Abgrenzung der Arten. Dtsch. ent. Z., 525-544, 705-736.
- Bôttcher, G. (1913), Die männlichen Begattungswerkzeuge der Arten bei dem Genus Sarcophaga Meigen. und ihre Bedeutung für die Abgrenzung der Arten. Dtsch. ent. Z., 1-16, 115-130, 239-254, 351-377.
- Brauer, F. & Bergenstamm, J.E., (1889), Die Zweiflügler des Keiserlichen zu Wien. IV. Vorarbeiten zu einer Monographie der Muscaria Schizometopa (exclusive Anthomyidae). Pars I. Denkschr. Akad. Wiss., Wien, 56:69-180.
- Brauer, F. & Bergenstamm, J.E., (1891), Die Zweiflügler des Keiserlichen zu Wien. V. Vorarbeiten zu einer Monographie der Muscaria Schizometopa (exclusive Anthomyidae). Pars II. Denkschr. Akad. Wiss., Wien, 58:39-446.
- Enderlein, G. (1928a), Klassification der Sarcophagiden. Sarcophagiden-Studien I. Arch. klassifik. phylogen. Ent., 1:1-56.
- Enderlein, G. (1928b), Sarcophgiden-Studien II. Konowia, 7:147-153.
- Enderlein, G. (1936), 22. Ordnung: Zweiflügler, Diptera. Die Tierwelt Mitteleuropas. Vol. 6, Insekten, Teil III, Abt. 16:1-259.
- FAN, ZI-DE (1965), Key to the common synanthropic flies of China. Academy of Sciences, Peking, XV + 1-330.
- FAN ZI-DE (ed.), 1992, Key to the common flies of China. Second dition. Shangai Institute of Entomology, Academia Sinica. 992p + 40 pl.
- Kano, R., Flield, G. & Shinonaga, S. (1967), Sarcophagidae (Insecta: Diptera). Fauna Japonica 7:1-168 + 41 pls.
- Lehrer, A.Z., (1974), Diptères myiasigènes de la superfamille Sarcophagidea de Roumanie. Acta Rer. Nat. Mus. Nat. Slov., Bratislava, 20:125-159.
- Lehrer, A.Z., (2000a). Le système taxonomique des Sarcophaginae afrotropicales (Diptera, Sarcophagidae). Entomologica, Bari, 34:41-63.
- Lehrer, A.Z. (2000b), La structure de l’abdomen des Sarcophaginae (Diptera, Sarcophagidae). Entomologica, Bari, 34:153-169.
- Lehrer, A.Z., (2003a), Revision du genre Wohlfahrtia Brauer & Bergenstamm, 1889 de la faune d’Israël (Diptera, Sarcophagidae). Entomol. Croat., 7, nr. 1-2: 15-33.
- Lehrer, A.Z., (2003b), Sarcophaginae de l’Afrique (Insecta, Diptera, Sarcophagidae). Entomologica, Bari, 37 :5-528.
- Lehrer, A.Z., (2005), Nouveaux Sarcophagides afrotropicaux et orientaux (Diptera, Sarcophagidae). Entomologica, Bari, 39,:5-59.
- Lehrer, A.Z., 2006, Sarcophaginae et Paramacronychiinae du Proche Orient (Insecta, Diptera, Sarcophagidae). Pensoft, 1-263.
- Lehrer, A.Z., 2008, Le statut taxonomique des espèces "Musca carnaria Linnaeus, 1758" et Sarcophaga carnaria Bottcher, 1912 (Diptera, (Diptera, Sarcophagidae). Fragmenta Dipterologica, 13:15-17 [1].
- Lehrer, A. Z. & Fromunda V., 1986, Le développement larvaire du diptèremyiasigène Wohlfahrtia magnifica (Schiner) (Diptera, Sarcophagidae). Bull. Ann. Soc. r. bege Ent., 122:129-136
- Lehrer A., Lehrer M. & Verstraeten C., 1988, Les myiases causées aux moutons de Roumanie par Wohlfahrtia magnifica (Schiner) (Diptera, Sarcophagidae). Ann. Méd. Vét., 132:475-481.
- Lehrer, A. Z. & Luciano P., 1980, Sarcophagides (Diptera) parasites de Porthetria dispar (L.) en Sardaigne et leur cartographie dans le reseau U.T.M. Studi Sassaresi, Sez. III, Ann. Fac. Agraria Univ. Sassari, 27:161- 173.
- Lehrer, A. Z. & Verstraeten C., 1991, Expansion parasitologique et géographique de Wohlfahrtia magnifica (Schiner) (Diptera, Sarcophagidae) en Roumanie. Bull. Rech. Agron. Gembloux, 26(4):563-567.
- Leonide, J. & Leonide J.-C. (1986), Les diptères sarcophagidés des orthoptères français – essai biotaxonomie. Université de Provence (Aix-en-Provence), 1-301.
- Meigen, J.W. (1826), Systematische Beschreibung der bekannten europäischen zweiflügeligen Insekte. Fünfter Teil, Schulz, Hamm., 1-412.Mihàlyi, F. (1975), Beschreibung vier neuer Sarcophagidae Arten aus Ungarn (Diptera), Acta zool. hung., 21:101-108.
- Mihàlyi, F., (1979b), Fémeslefgyek - Húslegyek. Calliphoridae – Sarcophagidae. Fauna Hung., 135:1-152.
- Pape, T. (1987), The Sarcophagidae (Diptera) of Fennoscandia and Denmark. Fauna ent. scand., 19:1-203.
- Pape, T., 1996, Catalogue of the Sarcophagidae of the World (Insecta:Diptera). Mem. on Entomology, Intern., vol. 8, 558 p.
- Povolny D. & Verves, YU.G. (1997), The flesh-flies of Central Europa (Insecta, Diptera, Sarcophagidae). Spixiana. Supplement, München, 24 :1-260.
- Rohdendorf, B.B. (1937), Fam. Sarcophagidae. (P. 1). Faune de l'URSS, 19:1-501 [In Russian with German summary].
- Salem, H.H. (1935), The Egyptian species of the genus Sarcophaga. Publ. Egypt. Univ. Fac. Med., 5:1-61.
- Seguy, E. (1941), Études sur les mouches parasites. 2. Calliphoridae, calliphorines (suite), sarcophagines et rhinophorines de l'Europe occidentale et méridionale, Encycl. ent. (Ser. A), 21:1-436.
- Senior-White, R.A., Aubertin, D. & Smart, J. (1940), Diptera. Family Calliphoridae. The fauna of British India, including the remainder of the Oriental Region. Vol. VI. London, 1-288.
- Thompson, F.C. & Pont, A.C., 1993, Systematic Database of Musca Names (Diptera). These Zoologicae, 20, 221 p.
- Verves, YU. G. (1982), 64h. Sarcophaginae. Die Fliegen der plaearktischen Region, Stuttgart, Bd. 11, Lf. 327:235-296.
- Verves, YU. G. (1985), 64h. Sarcophaginae. Die Fliegen der plaearktischen Region, Stuttgart, Bd. 11, Lf. 330:297-440.
- Zumpt, F. (1972), Calliphoridae (Diptera Cyclorhapha). Part IV. Sarcophaginae. Explor. Parc nat. Albert, Miss. G.F. de Witte (1933-1935), 101:1-264.

















































