Rétine - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
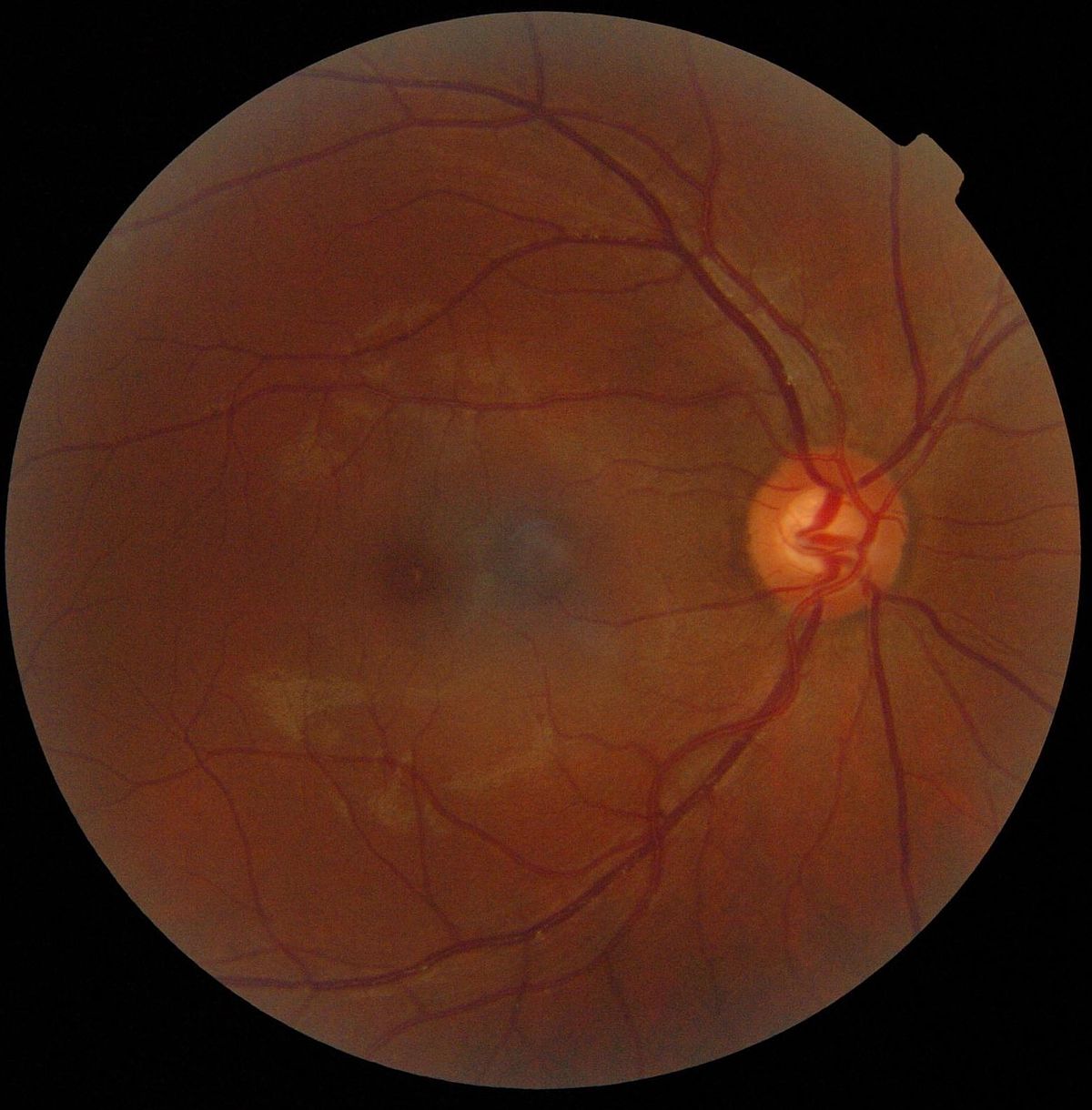
La rétine est l'organe sensible de la vision. D'origine diencéphalique, elle est une mince membrane pluri-stratifiée d'environ 0,5 mm d'épaisseur couvrant environ 75 % de la face interne du globe oculaire et intercalée entre l'humeur vitrée et l'épithélium pigmentaire sous-choroïdal. Sa partie sensible à la lumière se compose de photorécepteurs: environ 5 millions de cônes (vision diurne et colorée) et ~120 millions de bâtonnets (vision crépusculaire et nocturne en noir et blanc), qui captent les signaux lumineux (photons) et les transforment en signaux électro-chimiques. Elle se compose aussi de neurones qui à leur tour intègrent ces signaux chimiques (neurotransmetteurs) en signaux électriques à l'origine de potentiels d'action mais aussi de cellules gliales. Ces potentiels d'action générés par les cellules ganglionnaires seront acheminés par les nerfs optiques vers l'encéphale par les corps géniculés latéraux (relais thalamiques qui se projettent par les radiations optiques vers la scissure calcarine du lobe occipital, hypothalamus, colliculi supérieurs, noyaux du tractus optique accessoire). La rétine est vascularisée par l'artère et la veine centrales dites de la rétine.
Quelques zones particulières de la rétine
- la macula : région centrale de la rétine, située proche de l'axe optique;
- la fovéa : région centrale de la macula où se concentrent les cônes;
- la papille : région d'émergence du nerf optique et dépourvue de photorécepteurs.
De la transduction au potentiel d'action: schéma général
Les photorécepteurs
Il existe deux types de photorécepteurs qui transforment, ou transduisent, le signal lumineux (photon) en signaux électrique (potentiels récepteurs) puis chimique (neurotransmetteur = glutamate): les cônes pour la vision diurne et colorée dont on distingue trois sous-types L, M et S, en fonction de leur sensibilité spectrale, et les bâtonnets pour la vision crépusculaire et nocturne en "noir et blanc".
Histologiquement, les photorécepteurs se présentent avec un péricaryon autour du noyau cellulaire dont le pôle supérieur est surmonté successivement par un segment interne renfermant principalement ergastoplasme et mitochondries, puis un segment externe lequel est constitué d'un empilement de citernes membranaires élaborées dans le segment interne pour les bâtonnets, et de replis membranaires se surface décroissante pour les cônes. Ces citernes et replis membranaires renferment les pigments visuels. Au pole inférieur du péricaryon, des évaginations cytoplasmiques, ou pédicule, définissent des terminaisons synaptiques avec les cellules horizontales et les cellules bipolaires. Ces synapses revêtent un aspect particulier et sont dites en ruban ("ribbon synapses"). Elles se composent d'invaginations du pédicule où se logent dendrites de deux cellules horizontales et de deux à cinq cellules bipolaires. en regard, au sein du compartiment intracellulaire, se trouve un densité présynaptique autour de laquelle s'accumulent des vésicules emplies de Glutamate.
La sensibilité à la lumière: les pigments visuels
Chaque photorécepteur synthétise dans son segment interne au-dessus du péricaryon, et stocke dans la membrane plasmique de son segment externe des pigments composés d'une protéine porteuse qui détermine la sensibilité spectrale, et qui est couplée à une molécule chromophore qui capte les photons. Dans le cas des bâtonnets, le pigment correspond à la rhodopsine constituée par une protéine: l'opsine, et le chromophore : 11-cis rétinal, aldéhyde de la vitamine A. Pour les cônes, les pigments sont: cyanolabe (S), chlorolabe (M), et érythrolabe (L), respectivement sensibles à la lumière de longueur d'onde maximale : 420 nm (bleu), 530nm (vert) et 560 nm (rouge).
Cônes (C) et bâtonnets (B)s'opposent quant à la propriétés de leur pigments visuels: sensibilité forte (B) et faible (C), acuité forte (C) et faible (B), et adaptation forte (B) et faible (C).
La transduction
L'absorption d'un photon par le chromophore entraîne des modifications électro-chimiques transitoires des photorécepteurs qui affectent sa polarité, et la libération de Glu à son pôle basal.
Mécanismes biochimiques (exemple des bâtonnets):
Au repos (dans l'obscurité), les bâtonnets apparaissent spontanément dépolarisés (-40 mV)en raison de l'existence d'un flux rentrant de cations (sodium et calcium, et dans une moindre mesure (5%) magnésium et potassium). Ce flux est lié à l'ouverture de conductances transmembranaires sous l'action du GMPc (guanylate monoophosphate cyclique) synthétisé à partir de l'action de la guanylate-cyclase sur le GTP (guanylate triphosphate). Le flux de potassium active un canal antiport: le potassium entrant est excrété en même temps que du sodium est pompé à dans le milieu extracellulaire ce qui contribue à accroître le flux entrant global de cet ion.
L'illumination de la rétine provoque la capture de photon par la rhodopsine. Il s'ensuit une modification géométrique du chromophore (photo-isomérisation) qui passe du 11-cis rétinal en 11-trans-rétinal. Ce phénomène se produit au sein de la membrane des citernes des segments externes.
Cette photo-isomérisation conduit à l'activation d'une protéine également membranaire: la transducine dont une sous-unité alpha se détache et se recombine transitoirement à une molécule de GTP, et à une enzyme jusqu'alors inactive: la phosphodiestérase. Le complexe résultant concourt à activer la phosphodiestérase laquelle, à son tour, hydrolyse et donc inactive le GMPc en GMP. Or, le GMpc est responsable de l'ouverture des canaux cationiques à l'origine de la dépolarisation. Donc, la diminution de la concentration de GMPc entraîne une fermeture de ces canaux. La diminution du flux entrant de sodium repolarise alors la membrane.
Cette repolarisation qui se propage du segment externe à l'ensemble du photorécepteur se traduit au niveau des terminaisons synaptiques par une réduction de la libération de Glutamate dans la fente synaptique.
L'ensemble de ce mécanisme électro-chimique subit un amplification qui permet notamment de contre-balancer le bruit thermique (thermoisomérisation): 1 photon active 100 transducines qui activent 1000 phosphodiestérases.
Cascades d'activations/inhibitions sur les réseaux radiaires
Au repos dans l'obscurité, les photorécepteurs se trouvent spontanément dépolarisés, et libèrent à leur base un neurotransmetteur : le glutamate (Glu). Glu diffuse dans la fente synaptique vers les cellules bipolaires ON qu'il inhibe en se fixant sur des récepteurs métabiotropes APB, et vers les cellules bipolaires OFF, qu'il active en se fixant sur des récepteurs ionotropes. Il s'ensuit que seule les cellules ganglionnaire OFF ainsi activées par les cellules bipolaires OFF émettront des potentiels d'action au sein du nerf optique.
En présence de lumière dont l'intensité recrutera soit les cônes soit les bâtonnets, l'absorption des photons au niveau des segments externes des photorécepteurs se solde par une dépolarisation cellulaire et corrélativement par une diminution de l'excrétion de Glu. Il en résulte une activation des cellules bipolaires ON par désinhibition (et donc des cellules ganglionnaires ON associées), et une inhibition des cellules bipolaires OFF.
Champs récepteurs des cellules ganglionnaires: antagonisme centre/périphérie
L'activation d'une cellule ganglionnaire dépend de l'illumination d'une région circulaire de la rétine et son inhibition d'une région annulaire circonscrivant la précédente région. Ces deux régions concentriques déterminent le champ récepteur à centre ON (activateur) et à périphèrie OFF (inhibitrice). La configuration inverse comportant un centre OFF et une périphérie ON existe également. Dans le premier cas, l'illumination du pourtour OFF conduit à l'inhibition du récepteur, et celle simultanée des centre ON et pourtour OFF conduit à une activation décroissante en fonction de l'étendue de l'illumination du pourtour. En conséquence, l'activité électrique finale des cellules ganglionnaires traduit le contraste lumineux, estimé grâce à l'antagonisme centre/périphérie de leurs champs récepteurs.
Le centre ON se compose des circuits radiaires incluant successivement: photorécepteurs, cellules bipolaires (et amacrines AII pour les bâtonnets) et cellules ganglionnaires. La périphérie OFF dépend de la présence des cellules horizontales connectées aux photorécepteurs situées dans cette zone. En effet, l'illumination des photorécepteurs de la zone OFF entraîne l'activation des cellules horizontales sous-jacentes, couplées entre elles par des jonctions communicantes. Ces cellules horizontales s'hyperpolarisent et libèrent alors un neurotransmetteur: le GABA, qui inhibe les photorécepteurs de la zone ON. Les cellules horizontales sont ainsi responsables d'une inhibition latérale dont l'extension spatiale dépend du couplage des cellules horizontales entre elles par des jonctions communicantes qui propagent l'hyperpolarisation. Les cellules plexiformes contrôlent cette extension spatiale en découplant les cellules horizontales. Les cellules horizontales H1 pourvues d'un très long axone se connectent aux bâtonnets, et aux L et M cônes, alors que les cellules horizontales H2 dépourvues d'un tel axone se connectent aux cônes majoritairement de type S.

















































