Mitochondrie - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
Une mitochondrie (du grec mitos, fil et chondros, grain) est un organite à l'intérieur d'une cellule eucaryote, dont la taille est de l'ordre du micromètre. Son rôle physiologique est primordial, puisque c'est dans les mitochondries que l'énergie fournie par les molécules organiques est récupérée sous forme d'ATP (énergie contenue dans la liaison phosphate-phosphate), la source principale d'énergie pour la cellule eucaryote, par le processus de phosphorylation oxydative.
Historique
En 1857, Kölliker décrit les aspects de la mitochondrie dans le muscle. En 1890, Altmann décrit une technique de coloration des mitochondries qu'il appelle bioblastes et postule leur autonomie métabolique et génétique. En 1937, un scientifique allemand, Hans Adolf Krebs, construit un modèle qu’il appela « citric acid cycle ». Ce cycle a lieu dans la mitochondrie chez les eucaryotes. En 1940-43, Claude isole les mitochondries dans des cellules du foie. En 1948-50, Kennedy et Lehninger montrent que le cycle de Krebs, la bêta-oxydation et la phosphorylation oxydative ont lieu tous dans la mitochondrie. En 1978, Peter Mitchell obtient le Prix Nobel pour sa théorie chimiosmotique. En 1981, Anderson et son équipe découvrent la structure génétique de l’ADN mitochondrial humain. Finalement, Boyer et Walker, eux aussi, obtiennent le Prix Nobel pour leurs études sur la structure et le fonctionnement de l'ATP synthétase.
Origine
Une mitochondrie ne peut provenir que de la croissance et de la division d'une autre mitochondrie déjà existante. Normalement, avant la division cellulaire, la mitochondrie double sa masse puis se scinde en deux. Elles sont aussi capables de fusionner entre elles. Cette division débute par l'apparition d'un sillon de division sur la membrane interne. Elle a lieu pendant toute l'interphase et nécessite l'intervention de la protéine DRP1 (voisine de la dynamine). La réplication de l'ADN mitochondrial n'est pas limitée à la phase S du cycle cellulaire. Le nombre de mitochondries par cellule est régulé par l'activité cellulaire. Par exemple, une cellule musculaire au repos contient 5 à 10 fois moins de mitochondries qu'une cellule musculaire activée en permanence.
Le fait que la mitochondrie possède son ADN propre, comme les chloroplastes, indique une origine exogène : il est maintenant admis que les mitochondries proviennent de l'endosymbiose d'une α-protéobactérie il y a environ 2 milliards d'années. La théorie endosymbiotique de l'origine des mitochondries, a été développée et argumentée par Lynn Margulis dès 1966, puis a été appuyée par la découverte de l'ADN spécifique des mitochondries en 1980. Il semble qu'au cours de l'évolution l'ADN originel de la bactérie ait subi diverses évolutions, perdu un grand nombres de gènes, parfois transféré dans l'ADN de la cellule hôte. Parallèlement à ce report de la synthèse de certaines protéines vers l'hôte, ce dernier a développé un arsenal de translocases, enzymes permettant le transfert de ces protéines vers la matrice mitochondriale.
Structure

1 : membrane interne.
2 : membrane externe.
3 : espace inter-membranaire.
4 : matrice.
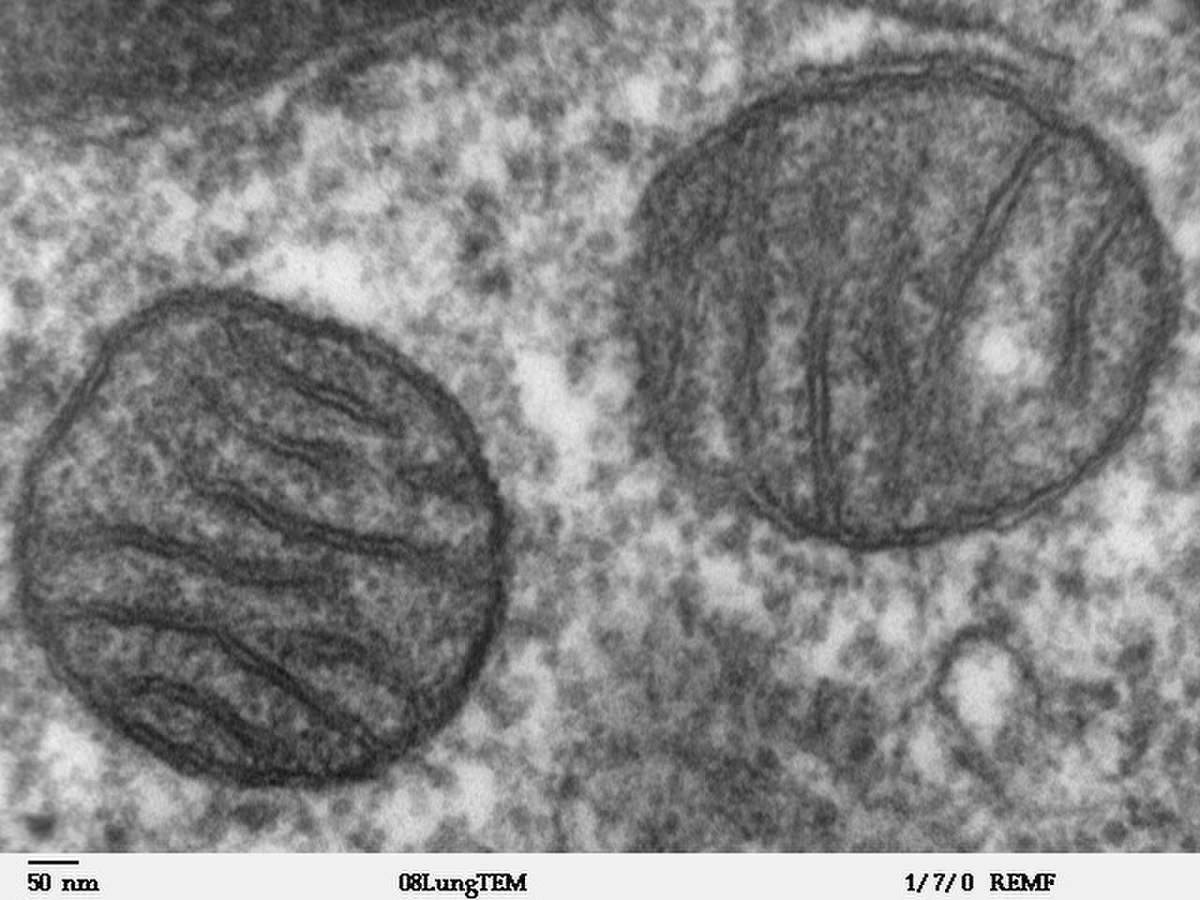
Les mitochondries ont une dimension de 1-2 à 10 μm de long et de 0,5 à 1 μm de large. Elles se composent de 2 membranes mitochondriales, une externe et une interne, qui délimitent trois milieux : le milieu extra-mitochondrial (cytoplasme de la cellule), l'espace inter-membranaire et la matrice. Chacune est de l'ordre des 6 nm et l'espace intermembranaire est de 7 nm.
- La membrane externe est formée de 60 % de protéines et de 40 % de lipides polaires. Elle contient de nombreuses protéines appelées porines (VDAC) qui forment des canaux aqueux au travers de la membrane. La porine (protéine transmembranaire composée de 16 feuillets béta formant les canaux protéiques traversant la couche bimoléculaire de lipides) laisse passer toutes les molécules hydrophiles d'une masse moléculaire inférieure à 10 000 daltons (anions, cations, les acides gras, le pyruvate, les nucléotides le traversent). La membrane externe présente des complexes TOM constitués de plusieurs sous-unités protéiques dont des récepteurs et des canaux aqueux qui permettent l'entrée des protéines d'origine nucléaire dans la mitochondrie ou l'insertion de ces mêmes protéines dans la membrane externe.
- La membrane interne est beaucoup moins perméable que la membrane externe. Elle est composée de 75 % de protéines et de 25 % de lipides. Elle contient en quantité un phospholipide double, la cardiolipine, renfermant 4 acides gras rendant cette membrane imperméable aux ions. Les autres molécules doivent passer par un transporteur pour traverser la membrane interne. La membrane interne présente des complexes TIM 23, TIM 22, et OXA. Le TIM 23 permet l'entrée de protéines situées dans l'espace inter-membranaire dans la matrice mitochondriale et dans la membrane interne. Le TIM 22 permet l'insertion des protéines dans la membrane interne et notamment des protéines à plusieurs domaines transmembranaires. Le complexe OXA permet la sortie de la matrice pour certaines protéines d'origine mitochondriale.
La membrane interne forme des invaginations qui apparaissent sous forme de crêtes ou replis au microscope électronique. Ces crêtes augmentent la surface de la membrane et donc de capacité de phosphorylation oxydative. Grâce à cette caractéristique on peut déduire que si une mitochondrie possède beaucoup de crêtes c'est que la cellule a besoin d'une grande quantité d'énergie et donc elle pourra produire plus d'ATP (cellule en activité). On retrouve également à son niveau des protéines de transport spécifiques pour les petites molécules utilisées par la matrice, les enzymes de la chaîne respiratoire, l'ATP-synthase ou complexe F0-F1 visible au microscope électronique sous forme de protubérance interne.

















































