Membrane plasmique - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
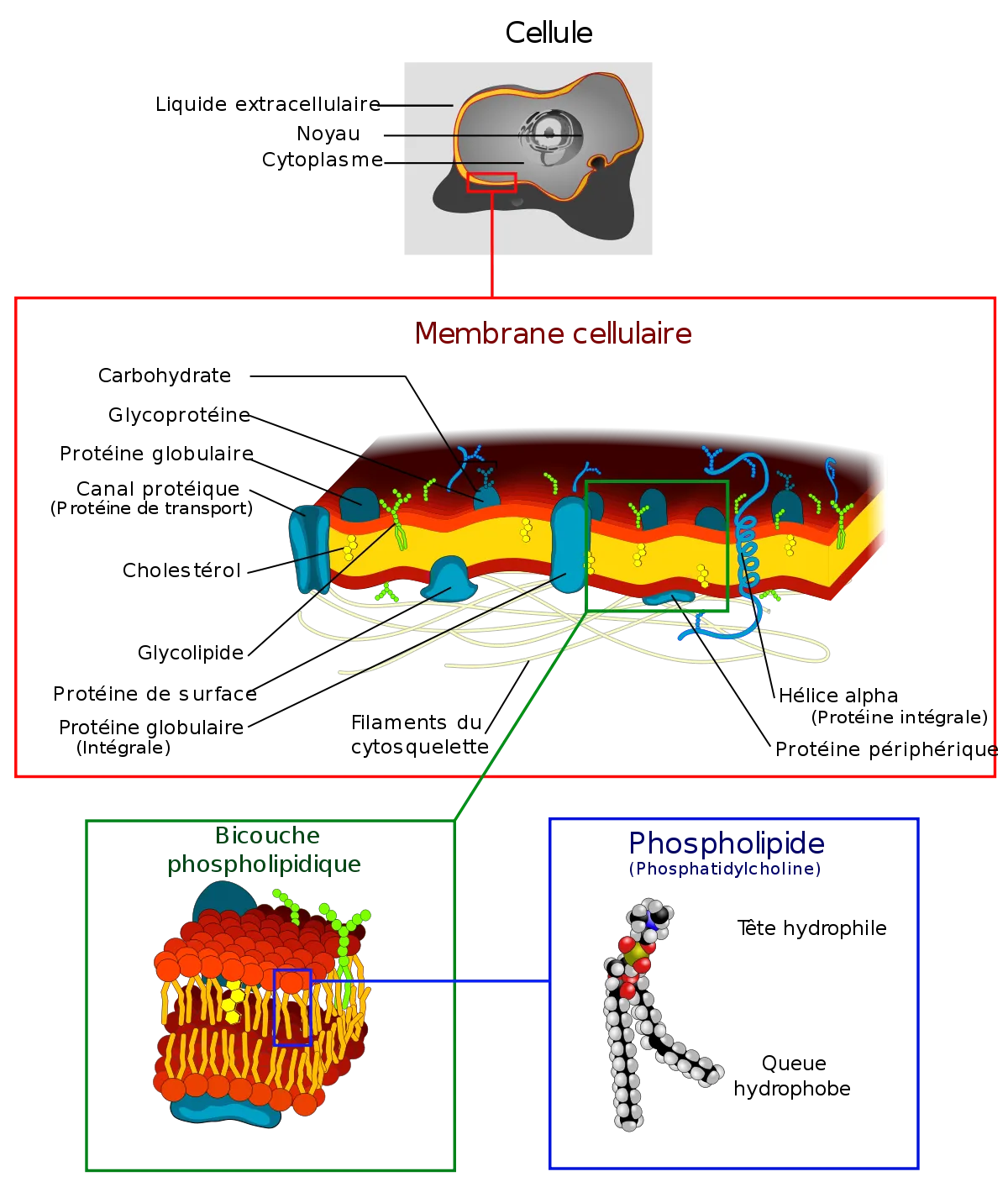
La membrane plasmique, ou plasmalemme, est la membrane qui délimite une cellule, elle sépare le cytoplasme du milieu extérieur. On distingue des sous-types de membranes, dont :
- sarcolemme (membrane de la cellule musculaire)
- névrilemme (membrane de la cellule nerveuse)
Description
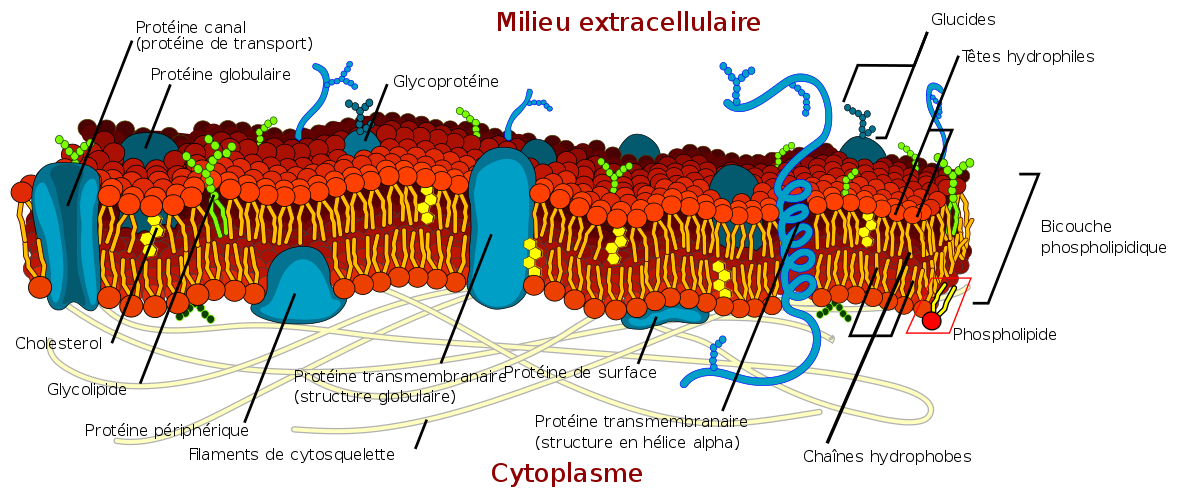
La membrane plasmique forme une pellicule continue de 5 à 10 nm d’épaisseur délimitant le cytoplasme du milieu extracellulaire. Ce micro-écotone est composée de phospholipides, de protéines (intracellulaires, extracellulaires ou insérées dans la double couche phospholipidique) et de molécules de cholestérol.
Grâce à une perméabilité très sélective, elle joue un double rôle de protection et de contrôle des échanges entre les milieux intracellulaire et extracellulaire.
Elle est à distinguer des endomembranes qui délimitent les organites.
Les glucides
Les sucres fixés sont des polysaccharides ou des oligosaccharides.
Ils sont toujours liés soit à des protéines (avec lesquels ils forment des glycoprotéines), soit à des lipides en formant alors des glycolipides. Les sucres les plus abondants sont des glycolipides fixés aux sphingolipides (et donc insérés dans les radeaux lipidiques).
Ils sont tous localisés à la face extracellulaire de la membrane, alors qu'à l'intérieur de la cellule où ils sont synthétisés, quand ils sont fixés, ils le sont sur les parois internes des cavités endomembranaires (organites à membranes).
Composition
Deux feuillets sont accolés ;
- l'hémimembrane E (« E » pour exoplasmique) ; C'est la couche externe de la membrane.
Ce feuillet est parfois lui-même recouvert d'une structure finement fibrillaire dite glycocalix ou cell-coat.
Sur cette face on trouve aussi fréquemment des glucides ou dérivés de glucides (sous forme d'acide sialique par exemple), dont toutes les fonctions ne sont pas encore comprises ; - l'hémi-membrane P, (« P » pour protoplasmique) côté interne, éventuellement plus ou moins liée à d'autres membranes intra-cellulaires.
Ces feuillets sont formés de phospholipides et de glycolipides (uniquement du côté extracellulaire) qui se font face par leur pôle hydrophobe.
Entre les molécules phospholipides se trouvent des molécules dites intrinsèques et du cholestérol (uniquement chez les eucaryotes et dans le monde animal, des phytostérols jouant un peu le même rôle chez les végétaux).
Sur les faces externe et interne, constituées par les pôles hydrophiles des phospholipides membranaires, se fixent des protéines extrinsèques sur lesquelles s’accroche le cytosquelette ou les molécules de la substance (matrice) extracellulaire.
- Cet assemblage non-covalent de lipides et de protéines, forme une structure souple permettant à presque tous ses éléments - dans une certaine mesure - de se déplacer latéralement. Cette « fluidité membranaire » sera d'autant plus importante que les acides gras constituant les lipides membranaires sont courts et insaturés.
- Des protéines spéciales assurent la perméabilité sélective de la membrane (pores), la reconnaissance de signaux chimiques ou portent des marqueurs (antigènes du CMH…).
Les pores sont les portes de la membranes. Ce sont des édifices macromoléculaires de nature protéique ou glycoprotéique, parfois très complexe. Ils sont insérés dans la bicouche et contrôlent les échanges entre milieu intra et extra cellulaire (avec des récepteurs, transporteurs).
La membrane est asymétrique et parfois irrégulière, en raison de
- différences de présence de lipides entre ses deux feuillets,
- différence de nature et composition en sucres,
- différence dans le mode d’insertion des protéines,
- différence de composition biochimique (d'un type cellulaire à l’autre, d'un endroit de la membrane à un autre ; aux niveaux apical ou basal par exemple). Ces différences évoluent parfois rapidement dans le temps.
Les lipides
Ce sont des constituants essentiels du « bi-couches » membranaire (environ 50 % du poids sec de la membrane) dont la cohérence est assurée par les oppositions de pôle hydrophile et hydrophobe (les lipides sont amphipatiques).
Deux types lipidiques sont plus abondants :
- le cholestérol ; bien que presque absent des membranes des compartiments intracellulaires, il est essentiel dans la membrane plasmique ; Avec 15 à 20 % du poids sec de la membrane (et jusqu'à 30 % dans la membrane des érythrocytes), il y est en moyenne numériquement autant présent que les lipides (en nombre de molécules) ; Ce cholestérol forme un film étanche supplémentaire disposé entre les lipides membranaires, qu'il consolide sans la rigidifier. Il bloque toutes les molécules hydrophiles qui pourraient accidentellement s'insinuer dans le film lipidique moins cohérent ;
- les phospholipides, répartis en deux familles :
- phosphoglycérides. Au nombre de trois (chez les mammifères), ces dérivés du glycérol sont
- la sphingomyéline (2;l*m0%), molécule polaire, dérivée de la sphingosine, avec une chaîne d'acides gras liée par un phosphate. Cette molécule contient de la choline, isolant électrique, composant principal de la gaine de myéline des axones.
- d’autres phospholipides, moins abondants, mais essentiels pour les transmission de signaux, dont
- - des dérivés de l'inositol, qui sont :
- - des Glycosyl-Phosphatidyl-Inositol (GPI ou glycosil phosphatidyle inositol), qu'on ne trouve que dans le feuillet externe. Ce GPI permet l' ancrage de certaines glycoprotéines au feuillet externe de la membrane plasmique ;
- - divers autres dérivés présents sur le feuillet interne de la membrane (rôle majeur dans la transmission du signal)
La partie polaire : Elle forme un film souple composée d'un plan de phosphate estérifié par une molécule polaire (éthanolamine, choline, sérine) et relié à une molécule hydrophobe (sphingosine : alcool aminé) ou à un phosphoglycéride (en l'occurrence du glycérol, qui est également une molécule polaire).
La partie hydrophobe : C'est un film d'acides gras (AG), disposés de telle sorte qu'ils ne forment pas de motif de type cristallin, ce qui garantit à la membrane un bon compromis entre cohérence, résistance (forces de Van der Waals) et souplesse, avec des propriétés électriques intéressantes, la membrane isolant en quelque sorte la cellule et ses organites.
Les protéines
- Elles représentent l'autre moitié du poids sec de la membrane, mais occupent un volume plus important que les lipides qui sont sont plus petits que les protéines (50 molécules de lipides = 1 molécule de protéine en masse).
- Le type de protéine varie selon le type de cellule et ses fonctions. De même pour leur quantité : très abondantes dans la membrane interne de la mitochondrie ou dans la membrane du réticulum endoplasmique (70 % de la masse membranaire), abondante dans la membrane des hématies, hépatocytes ou des érythrocytes (50 % de la masse membranaire), elles sont plus éparses dans la membrane des cellules nerveuses (qui doivent être électriquement isolées, d'où l'abondance (20 % du poids sec de la membrane) de myéline).
- Elles sont divisées en deux classes :
- - protéines dites « extrinsèques » (ou périphériques) ; Elles sont insérées dans la bicouche lipidique, mais par des liaisons de faible énergie (Force de van der Waals) ; une simple variation de pH ou des conditions ioniques suffit à les en détacher ;
- - protéines dites « intrinsèques » ; transmembranaires ou solidement greffées par une liaison covalente à l'un ou l'autre des feuillets de la bicouche (à l’aide d’un acide gras ou par un GPI) ; Elles ne se détachent pas de la membrane ; le seul moyen de les extraire est de détruire la bicouche lipidique membranaire en utilisant un détergent puissant.

















































