Méiose - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
Il existe deux types de divisions cellulaires dans le monde vivant : la mitose qui assure la naissance de cellules identiques à la cellule mère lors de la multiplication asexuée et la méiose qui aboutit à la production de cellules sexuelles ou gamètes pour la reproduction.
Chez les animaux, la méiose est un processus se déroulant durant la gamétogénèse (spermatogenèse ou ovogenèse), c'est-à-dire durant l'élaboration des gamètes (les spermatozoïdes chez le mâle et les ovules chez la femelle) chez les espèces dites diploïdes.
Chez les végétaux, la méiose produit des spores, qui par mitose donneront une génération haploïde (le pollen, le pied feuillé des mousses, etc.) Elle donne des cellules haploïdes (cellules contenant n chromosomes) à partir de cellules diploïdes (cellule contenant 2n chromosomes - chez l'homme, une cellule normale contient 2n = 46 chromosomes (donc 23 paires) alors qu'un gamète contient n = 23 chromosomes au cours de deux divisions. Chez les espèces haploïdes (comme la Sordaria macrospora), la méiose intervient après la fécondation pour diviser la cellule-œuf (avec 2n chromosomes). Mais en plus de ce rôle de division, la méiose a un rôle important dans le brassage génétique (mélange des gènes) et ce, grâce à deux mécanismes de brassage : le brassage interchromosomique et le brassage intrachromosomique.
Ainsi, durant la méiose, la quantité d'ADN au sein de la cellule évolue au cours du temps.
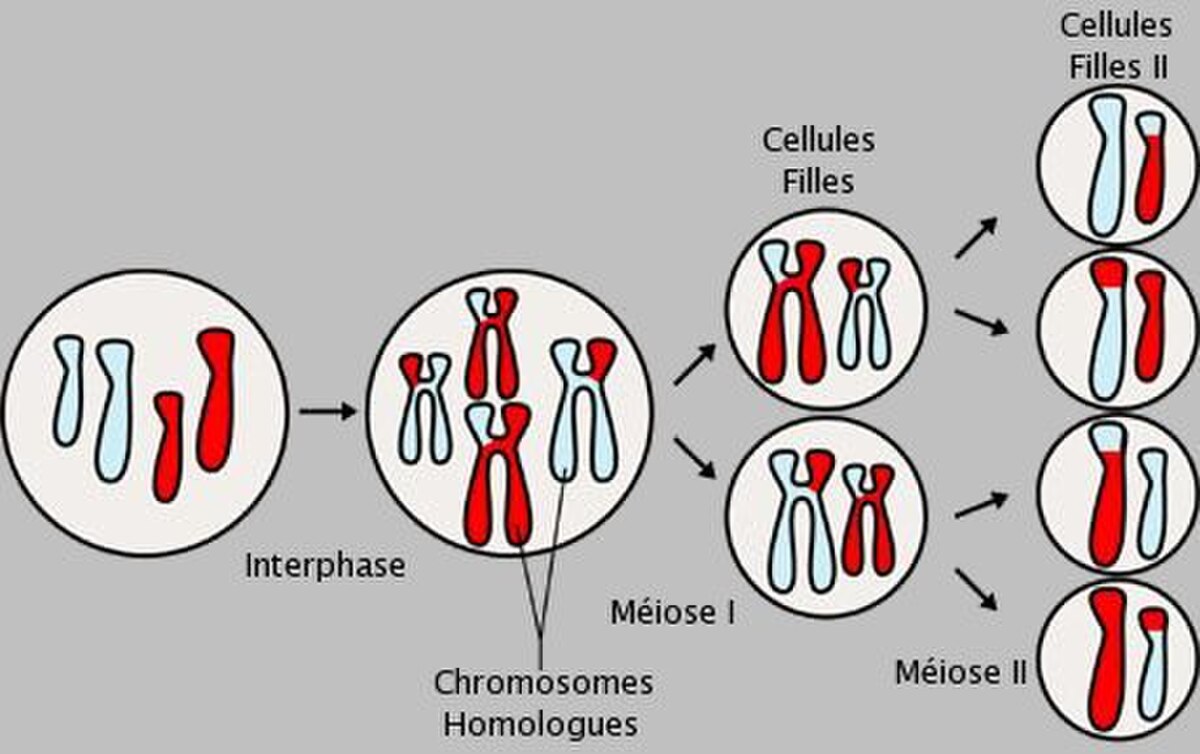
Chaque cellule va donc séparer son patrimoine génétique (contenu dans des chromosomes) en deux afin de ne transmettre que la moitié de ses gènes aux cellules filles.
Elle se déroule en plusieurs étapes formant un ensemble de deux divisions cellulaires, successives et inséparables.
Schéma
Deuxième division : méiose équationnelle
La méiose équationnelle consiste en une simple mitose, à la différence près du nombre de chromosomes qui est de n.
- Prophase II : Phase identique à la prophase I mais brève car les chromosomes sont restés compactés.
- Métaphase II : Les chromosomes se placent sur la plaque équatoriale.
- Anaphase II : Les chromatides de chaque chromosome se séparent et migrent vers des pôles opposés de la cellule.
- Télophase II : La cellule se sépare en deux, formant ainsi quatre cellules à n chromosomes à une chromatide.
À l'issue de cette deuxième division de la méiose on passe de 2 cellules mères à n chromosomes bichromatidiens à 4 cellules filles à n chromosomes monochromatidiens.
Première division : méiose réductionnelle
Prophase I
La prophase I est divisée en cinq étapes qui correspondent à cinq états caractéristiques de la chromatine : leptotène, zygotène, pachytène, diplotène et diacinèse.
- Leptotène : Début de la condensation de la chromatine et attachement des télomères (extrémités des chromosomes) à l'enveloppe nucléaire.
- Zygotène : Début de l'appariement des chromosomes homologues (synapsis) par le complexe synaptonémal (ou synapton) et convergence des télomères. Le complexe synaptonémal est une structure complexe constituée d'un élément central, SYCP1 qui forme un homodimère, relié à deux éléments latéraux. Les élements latéraux sont en fait les cohésines SMC1, SMC3 formant un hétérodimère maintenu en place par hREC8 hHR21. Les cohésines se trouvent de part et d'autre par des filaments transverses, à celles-ci se lie la chromatine de chaque zone des chromosomes impliqués dans le phénomène ultérieur d'enjambement (ou crossing-over). Il y a organisation « en bouquet » des chromosomes. L'ensemble des deux chromosomes homologues s'appelle une tétrade (car 4 chromatides) ou un bivalent (car 2 chromosomes).
- Pachytène : Appariement strict des chromosomes homologues et apparition des nodules de recombinaison et de nodules tardifs qui permettent les enjambements (échanges entre chromatides homologues). Cette phase a une importance considérable dans le brassage chromosomique.
- Diplotène : Désynapsis (séparation des chromosomes homologues), mais les chromosomes restent attachés en plusieurs points au niveau desquels deux des quatre chromatides semblent s'entrecroiser (chiasma). Pour le bon déroulement de la méiose il en faut au minimum un par chromosome, en moyenne 2-3. Il y a décondensation de la chromatine et formation des grandes boucles permettant un fort taux de transcription. Cette étape de la prophase I peut durer plusieurs années chez l'ovocyte.
- Diacinèse : Recondensation de la chromatine et détachement des télomères de l'enveloppe nucléaire. Glissement des chiasmas vers les télomères (terminalisation des chiasmas). À la fin, il y a disparition de l'enveloppe nucléaire.
Métaphase I
Les paires de chromosomes homologues (bivalents) se placent de part et d'autre. Pour chaque bivalent, les centromères se placent de part et d'autre ainsi qu'à égale distance du plan équatorial. Leur orientation se fait de façon aléatoire : on appelle ce phénomène la « ségrégation indépendante ». Cette ségrégation permet un second degré de diversification des cellules-filles : le brassage interchromosomique.
Anaphase I
Chaque chromosome s'éloigne de son homologue et migre au pôle opposé, tiré par des microtubules kinétochoriens (microtubules accrochés à un kinétochore au niveau d'un centromère) dû à la dépolymérisation de tubuline. Il n'y a pas clivage des centromères, ceci est dû au fait que la séparase dégrade hREC8 (et donc la construction des cohésines) mais au centromère est inefficace étant donné que Sga1 y protège hREC8.
Télophase I
Les enveloppes nucléaires réapparaissent dans chaque cellules, il y a donc formation de deux cellules haploïdes à n chromosomes à deux chromatides (chromosomes bichromatidiens)(n chromosomes, 2n ADN). La cellule se divise en deux, grâce à un anneau contractile fait d'actine et de myosine.

















































