Magnoliophyta - Définition
La liste des auteurs de cet article est disponible ici.
La reproduction des Angiospermes
Les Angiospermes présentent le système reproductif le plus évolué des végétaux. Ce sont d'ailleurs les organes reproducteurs (fleurs et fruits) qui sont les caractéristiques les plus visibles du groupe.
La sexualité chez les Angiospermes
L'organisation des pièces florales
Les termes fleur et plante sont souvent confondus par les non botanistes. Or, il doit être noté que les fleurs ne constituent que la partie reproductive de certaines plantes — les plantes à fleur. La fleur est constituée de plusieurs pièces, dont les étamines (constituant la partie mâle), et le pistil (constituant la partie femelle). Une fleur hermaphrodite est une fleur possédant à la fois étamines et pistil.
La répartition des sexes chez les plantes à fleurs
Les Angiospermes sont à reproduction majoritairement sexuée, avec quelques cas de parthénogenèse (pissenlits).
La majorité (70%) sont hermaphrodites (les individus ont les deux sexes), avec en général des fleurs mixtes (voir Herkogamie). Cependant, chez d'autres espèces, les fleurs mâles et femelles sont séparées sur le même individu (espèces monoïques, telles que le noisetier et le maïs). D'autres combinaisons sont possibles (fleurs hermaphrodites et fleurs mâles sur le même pied par exemple).
Quelques espèces (environ 7%) sont à sexes séparés sur plante séparée (les individus sont mâles ou femelles, par exemple le houx, le kiwi, on parle d'espèce dioïque), voire fleur hermaphrodite et fleur mâle sur pieds différents (espèce gynodioïque). Enfin, on note des cycles reproductifs plus curieux, comme le cas du figuier.
Les individus ne sont pas nécessairement auto-fécondables, c'est-à-dire que la fécondation n'est pas toujours possible sur la même plante (nombreux cultivars de pommiers).
Cycle de vie des Angiospermes
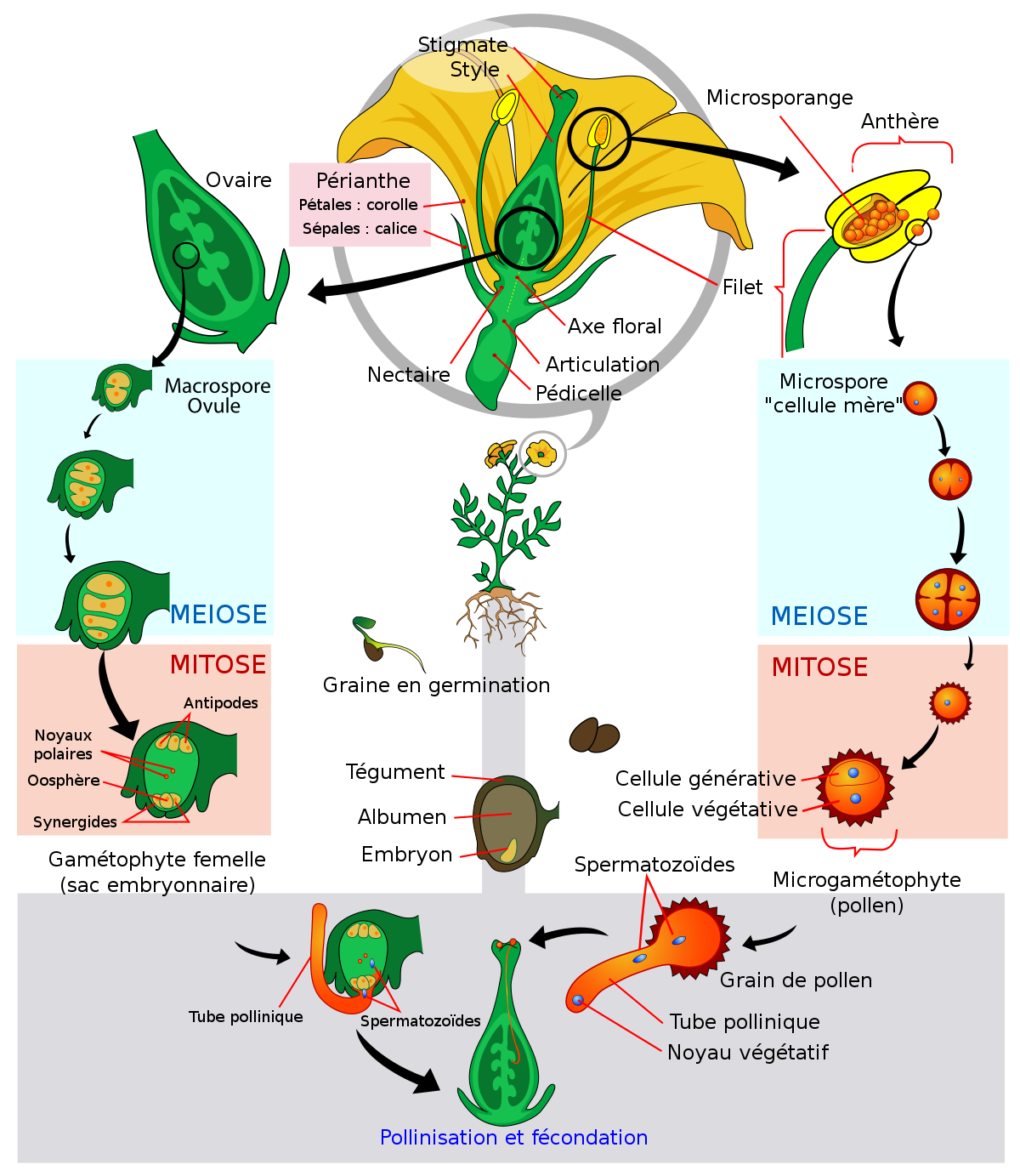
Comme toutes les plantes, les Angiospermes présentent une alternance entre deux états : l'état sporophytique et l'état gamétophytique. Cependant cette alternance fait partie d'un cycle très déséquilibré : la phase gamétophytique est très réduite dans l'espace et dans le temps, ce qui correspond à une meilleure adaptation au milieu de vie non aquatique des Angiospermes. En effet l'exposition au soleil augmentant les risques de mutations, une dominance de la phase sporophytique (au plus grand niveau de ploïdie) évite d'exprimer directement les mutations récessives. De plus le sporophyte (par exemple un arbre) est invariablement plus résistant chez les plantes.
Les gamétophytes mâle et femelle correspondent respectivement au grain de pollen et au sac embryonnaire chez les Angiospermes. On aurait donc tort de penser que le grain de pollen ou que le sac embryonnaire (lui-même contenu dans l'ovule, d'où la confusion) sont des gamètes : c'est en effet complètement faux puisqu'ils contiennent les gamètes ou les futurs gamètes.
Le grain de pollen
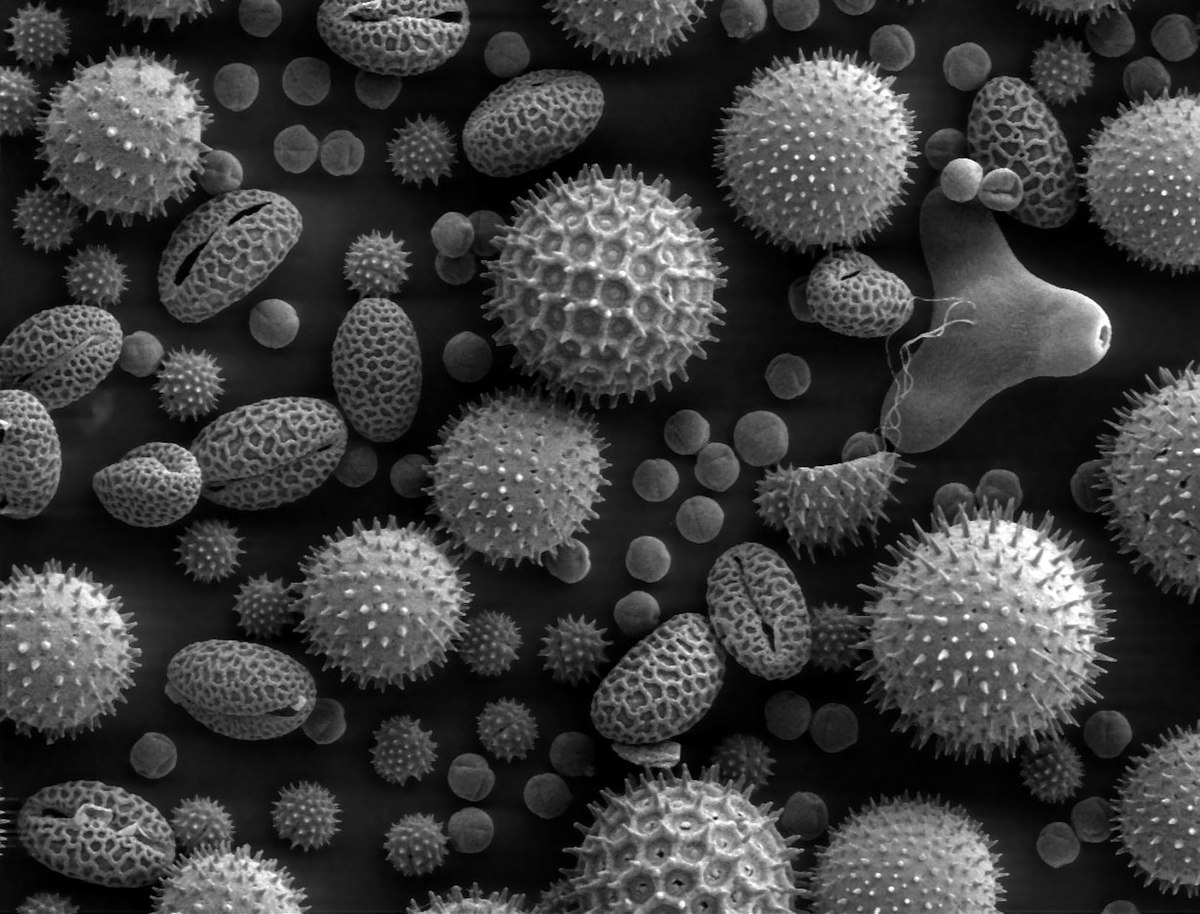
Généré par les anthères, le grain de pollen correspond au gamétophyte mâle. Il renferme deux à trois cellules et est entouré d'une double paroi formée du matériau le plus résistant du monde vivant: la sporopollénine qui le rend fossilisable. Cette paroi est constituée de deux couches non cellulaires: l'intine, interne qui est continue et secrétée par le gamétophyte et l'exine, externe, qui présente des apertures et qui est secrétée par le sporophyte. Dans le cas d'un pollen bicellulaire, le grain de pollen contient une cellule végétative (qui germera pour former le tube pollinique) et une cellule génératrice (qui correspond aux futures gamètes mâles, il s'agit d'une cellule n'ayant pas accompli sa deuxième division de méiose). Dans le cas du pollen tricellulaire, la cellule génératrice a déjà accompli sa deuxième division de méiose.
Le sac embryonnaire
Il peut être monosporé ou tétrasporé, c’est-à-dire provenir d'une seule mégaspore du sporophyte ou d'une tétrade de spores issues de la même méiose. C'est un tissu (non pas un gamète) gamétophytique (contenant le gamète femelle). Il peut présenter 7 ou 4 cellules. Dans le cas d'un sac embryonnaire à sept cellules, y sont présentes:
- l'oosphère, c’est-à-dire la cellule fécondable ou le gamète femelle.
- deux synergides, cellules adjuvantes de la fécondation qui forment un tore autour de l'oosphère et qui assumeront le rôle de sas pour faire transiter le gamète mâle. Ces cellules peuvent éventuellement générer un embryon non zygotique dans certains cas de reproduction asexuée.
- Trois antipodes situés à l'autre bout du sac embryonnaire.
- Une cellule centrale à deux noyaux (dits polaires) qui fusionneront avec le deuxième gamète mâle lors de la double fécondation pour éventuellement donner l'albumen.
Le sac embryonnaire renferme donc et protège le gamète femelle dont il favorisera la fécondation. Il est lui-même enfermé dans le nucelle à l'intérieur de l'ovule bitégumenté.
La pollinisation

Du fait de leur immobilité, la pollinisation d'un végétal terrestre par un autre géographiquement éloignée pose une difficulté. Si une possibilité est de s'en remettre au vent (pollinisation anémophile), les Angiospermes présentent la particularité que leur pollen est très souvent transporté par des espèces animales (pollinisation zoophile), principalement des insectes (pollinisation entomogame) mais aussi des chauve-souris et des marsupiaux (pollinisation cheiroptérogame, rencontrée chez le baobab) et des oiseaux (pollinisation ornithogame), qui se trouvent être attirés par la couleur, la forme, l'odeur ou le nectar des fleurs. Ce qui permet aux auteurs de dire Dans une fleur, les pétales participent également à la rencontre des gamètes. Et en effet l'organisation des pièces florales joue un rôle très particulier dans la fécondation, ainsi que nous allons développer un peu plus loin. La disparition des abeilles (syndrome d'effondrement des colonies d'abeilles) pourrait avoir des conséquences sur l'évolution de la biodiversité des plantes à fleurs dans les années à venir.
La fécondation
La fécondation se fait par germination du pollen sur le pistil, sans nécessité d'humidité comme dans le cas des fougères (caractère qu'ils partagent avec les autres Spermaphytes).
Chez les Angiospermes la fécondation présente trois caractéristiques essentielles:
- elle est siphonogame : le grain de pollen aperturé possédant deux ou trois cellules voit sa cellule végétative germer et former un tube pollinique qui acheminera, à l'aide de son cytosquelette, le gamète mâle (cas du pollen tricellulaire) où la cellule génératrice (cas du pollen bicellulaire) qui y effectuera sa seconde division de méiose. En ceci la siphonogamie correspond à un total affranchissement de l'eau pour la fécondation.
- elle est anisogame : le gamète femelle est de loin le plus massif des deux (le grain de pollen étant souvent très déshydraté, les cellules qu'il contient ont un espace cytoplasmique très réduit). Ceci implique que l'embryon possède systématiquement les caractères cytoplasmiques de sa mère. La transmission des gènes chloroplastiques et mitochondriaux (qui jouent un rôle important dans certains cas de résistance aux pesticides comme l'atrazine ou aussi dans des cas de stérilité mâle) n'est pas mendelienne. La mère transmet son cytoplasme en bloc et donc son génome mitochondrial et chloroplastique.
- elle est double : les gamètes mâles acheminés par le grain de pollen vont fusionner leurs noyaux (caryomixie) avec ceux de deux cellules différentes. Une première caryomixie entre gamète mâle et oosphère donnera l'embryon, une autre entre le second gamète mâle et les noyaux centraux de l'ovule conduira à une cellule triploïde qui formera l'albumen de la graine.
Voir aussi : le tube pollinique, le stigmate
La graine
Une fois fécondée, l'oosphère se développera en embryon. L'ovule contenant le sac embryonnaire qui la renfermait va donc former la graine (quant à l'ovaire, il formera le fruit vrai, l'ensemble graine et fruit étant appelé diaspore). On détermine trois catégories principales de graines en fonction de leur lieu de stockage des réserves:
- Les graines albuminées ou graines à albumen. Les deux produits de la double fécondation s'y développent ensemble: le zygote polarisé donne l'embryon et le noyau triploïde (3x) de la cellule centrale se divise très vite au sein de la même cellule (coenocyte) puis a lieu une étape de cloisonnement cellulaire pour former un tissu nourricier: l'albumen. La noix de coco par exemple présente un « lait » qui correspond en fait à la phase coenocytique de son développement. Observé au microscope, le « lait » de coco révèle qu'il est en fait un continuum de cytoplasme: c'est une cellule géante ayant un très grand nombre de noyaux. Une fois cellularisé, ce coenocyte formera le coprah.
exemples: Magnoliacées, Euphorbiacées (Hévéa, ricin, manioc), Ombellifères, Poacées (ou Graminées).
- Les graines exalbuminées, sans albumen. Les divisions du noyau triploïde central s'interrompent très vite mais l'embryon se développe normalement à partir du zygote. Il stockera ses réserves dans ses cotylédons. Exemples: Rosacées (pommier), Fabacées (ou Légumineuses), Composées, Juglandées (noyer).
- les graines à périsperme, sans albumen. Les divisions du noyau triploïde central s'interrompent aussi très rapidement. L'embryon ne stocke pas ses réserves lui-même, c'est le nucelle qui assumera cette fonction en devenant le périsperme. Exemples: Musacées (bananier), Nymphéacées, Chénopodiacées (betterave, épinard).
La graine peut être plus ou moins déshydratée. La longévité étant proportionnelle au degré de déshydratation, certaines graines (notamment pour les plantes des régions intertropicales sans saison défavorable) le sont peu (exemple: fève du cacaoier), d'autres le sont à des niveaux extrêmes (masse d'eau inférieure à 1% de la masse totale chez Nelumbo, le lotus d'Asie dont la longévité est de l'ordre du millier d'années.)
Le fruit
La dispersion de la progéniture
L'ensemble fruit et graine est appelé diaspore, le fruit est une structure uniquement ovarienne (c'est pourquoi on parle dans de nombreux cas de « faux fruits »,(et non pas la pomme , la fraise en faisant partie) tandis que la graine provient de l'ovule.
La graine et le fruit correspondent à une certaine forme de viviparité. L'embryon des Angiospermes se développe dans un premier temps protégé et alimenté par son parent femelle, puis est libéré, dans un état de vie ralentie (diapause), muni de protections et de réserves alimentaires (le fruit). Certains fruits présentent en plus des caractéristiques permettant la dispersion sur des distances importantes. On peut citer les aigrettes des pissenlits, qui font que le vent transporte les graines sur des dizaines de mètres. Citons aussi la mise à contribution de mammifères pour le transport (par accrochage dans les poils ou, moins important il faut l'admettre dans les chaussettes de campeurs). Un autre mode de dispersion s'observe quand des animaux consomment certains fruits : ils peuvent alors lâcher la graine et ses enveloppes non comestibles loin de son parent (avant ou après transit intestinal, selon). Il est par ailleurs clair qu'un mimétisme et une coévolution peuvent être en cause dans l'adaptation des plantes à fleurs à la faune capable de disperser leur progéniture. On connaît par exemple certaines graines dont la dormance est spécifiquement levée par des enzymes digestives d'animaux spécifiques (par exemple certains acacias africains).
Aussi : péricarpe, akène, drupe, fruit parthénocarpique, fruit charnu, fruit sec indéhiscent
Spécificité des conditions naturelles
Étant fixées et en milieu non aqueux, les Angiospermes ont à faire face à un très grand nombre de pressions sélectives. c'est en particulier pour cette raison qu'elles sont beaucoup plus diversifiées que les Thallophytes (algues) qui, elles, se développent au sein d'un milieu très peu changeant. De fait les Angiospermes représentent le sous-embranchement le plus diversifié des Chlorobiontes (lignée verte). Cette diversification passe nécessairement par une diversité génotypique. Ainsi la descendance d'une angiosperme en conditions sauvages a toujours tendance à ne pas être homogène, et donc à présenter une très large gamme de génotypes différents.
Pour entretenir cette diversité génétique, les plantes ont recours à l'allogamie (régime de reproduction sexuée chez les espèces dont les individus s'interfécondent systématiquement, par opposition à l'autogamie qui a été sélectionnée par l'Homme, nous verrons pour quelles raisons) qui rend incontournable la large diffusion du pollen. Une seule descendance compte ainsi un très grand nombre de génotypes différents qui conduiront à des phénotypes dont certains seront certainement très bien adaptés au milieu (les autres mourront). Les plus adaptés disposant ensuite de la reproduction asexuée pour se multiplier avant d'avoir atteint la maturité sexuelle. On connaît divers modes favorisant l'allogamie (s'opposant à l'autogamie).
- Les Systèmes d'(auto)Incompatibilité Sporophytiques et Gamétophytiques (SIS et SIG). Il s'agit de réactions de rejet de l'autopollen (pollen provenant du soi). On l'a vu, le grain de pollen est composé de deux à trois cellules et est entouré du sporoderme essentiellement composé du matériau le plus résistant chez le Vivant : la sporopollenine. ce sporoderme se compose de deux couches : l'intine et l'exine dont les origines sont très différentes. L'exine (qui présente les apertures) a été synthétisée par le sporophyte, et présente donc certaines molécules (RNAses et glycoprotéines) qui résultent de l'expression d'allèles sporophytiques. Ces protéines sont reconnues lorsque l'autopollen arrive au niveau du stigmate, et elles déclenchent l'autoincompatibilité Sporophytique (par reconnaissance de molécules synthétisées par le sporophyte). Ce rejet se rencontre dans le cas de pollen tricellulaire.
L'autoincompatibilité Gamétophytique repose sur le même principe, mais est basée sur la reconnaissance de substances synthétisées par le gamétophyte mâle (c’est-à-dire le grain de pollen chez les Angiospermes). Ces substances sont contenues dans l'intine et sont reconnues au moment de la germination du pollen qui est alors rejeté. Le SIS se rencontre chez des espèces à pollen bicellulaire.
- Les décalages de maturité. Chez certaines espèces les pièces mâles de la fleur sont matures avant les pièces femelles (protandrie) ou après (protogynie, plus rare), ce qui réduit considérablement le nombre d'autofécondations.
- Le dimorphisme floral : les espèces ayant développé ce système présentent deux types d'individus en proportions égales dans leur population. Des individus à fleurs longistylées (style long, stigmate au-dessus des étamines, petits grains de pollen et grosses papilles stigmatiques) et des individus à fleurs brévistylées (style court, étamines au-dessus du stigmate mais gros grains de pollen et petites papilles stigmatiques). Ce dimorphisme empêche l'autofécondation car dans les deux cas l'autopollen ne peut pas pénétrer le stigmate du style, soit parce qu'il est trop gros, soit parce que le style est trop haut.

















































