Kératine 8 - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
La kératine 8 est une protéine cytoplasmique qui constitue les filaments intermédiaires au niveau des tissus épithéliaux à simple couche.
Description
Trois constituants forment le cytosquelette, les microfilaments, les microtubules et les filaments intermédiaires. Les filaments intermédiaires sont codés par 67 gènes distincts et existent en six sous-types, qui sont des polymères en super hélice de différents types de protéines fibrillaires (kératines de type acide, kératines de type basique, vimentine et apparentés, neurofilaments, lamines ainsi que des protéines fibrillaires en rapport avec les cellules gliales).
La kératine 8 est une protéine cytoplasmique qui constitue les filaments intermédiaires au niveau des tissus épithéliaux à simple couche. Il s’agit d’une cytokératine de type II basique qui s’associe de façon non covalente en hétérodimère insoluble, avec la cytokératine 18 de type I acide. Chez l’humain, son gène est situé sur le chromosome 12 et est composé d’environ 9,8 kpb. La protéine encodée possède 483 acides aminés, a un poids moléculaire de 53,7 kDa et son point isoélectrique est situé à 5,6.
Modification post-transductionnelle
La kératine 8 possède trois sites majeurs de phosphorylation in vivo au niveau de ses régions globulaires. L’emplacement de ces sites renseigne grandement sur la spécificité d’action d’une telle modification. On suppose qu’en étant situé sur les extrémités de la protéine, la phosphorylation de ces sites entraînerait à la fois une réorganisation des filaments intermédiaires, un déplacement dans l’équilibre de la portion insoluble polymérisée vers l’état soluble dépolymérisée, une relocalisation de la protéine au niveau des différents organites, une association avec un membre de la famille de protéines 14-3-3 et serait associé à la protection contre différents stress physiologiques. Le premier, situé au niveau de la sérine 23 dans la portion N-terminal de la protéine, est un site de phosphorylation extrêmement conservé chez les kératines de type II suggérant qu’il a une fonction commune à toutes ces types de protéines. Le second site se situe au niveau de la sérine 431 dans la portion C-terminale de la protéine et a comme rôle d’augmenter la spécificité des autres sites de phosphorylation durant la mitose et l’exposition au facteur de croissance épidermique en association avec la réorganisation des filaments intermédiaires. Le troisième site se situe au niveau de la sérine 73 dans la portion N-terminale et joue un rôle significatif dans la réorganisation des filaments. Dans le foie, les hépatocytes n’expriment que la cytokératine 8 et son homologue, la cytokératine 18. Maintenant on comprend le caractère toxique conféré à l’alcool qui induit une hyperphosphorylation de ces sites ayant comme conséquence directe la désorganisation des filaments intermédiaires. Une telle désorganisation est à l’origine de lésions hépatiques retrouvées entre autres lors de cirrhose du foie alcoolique.
Structure moléculaire
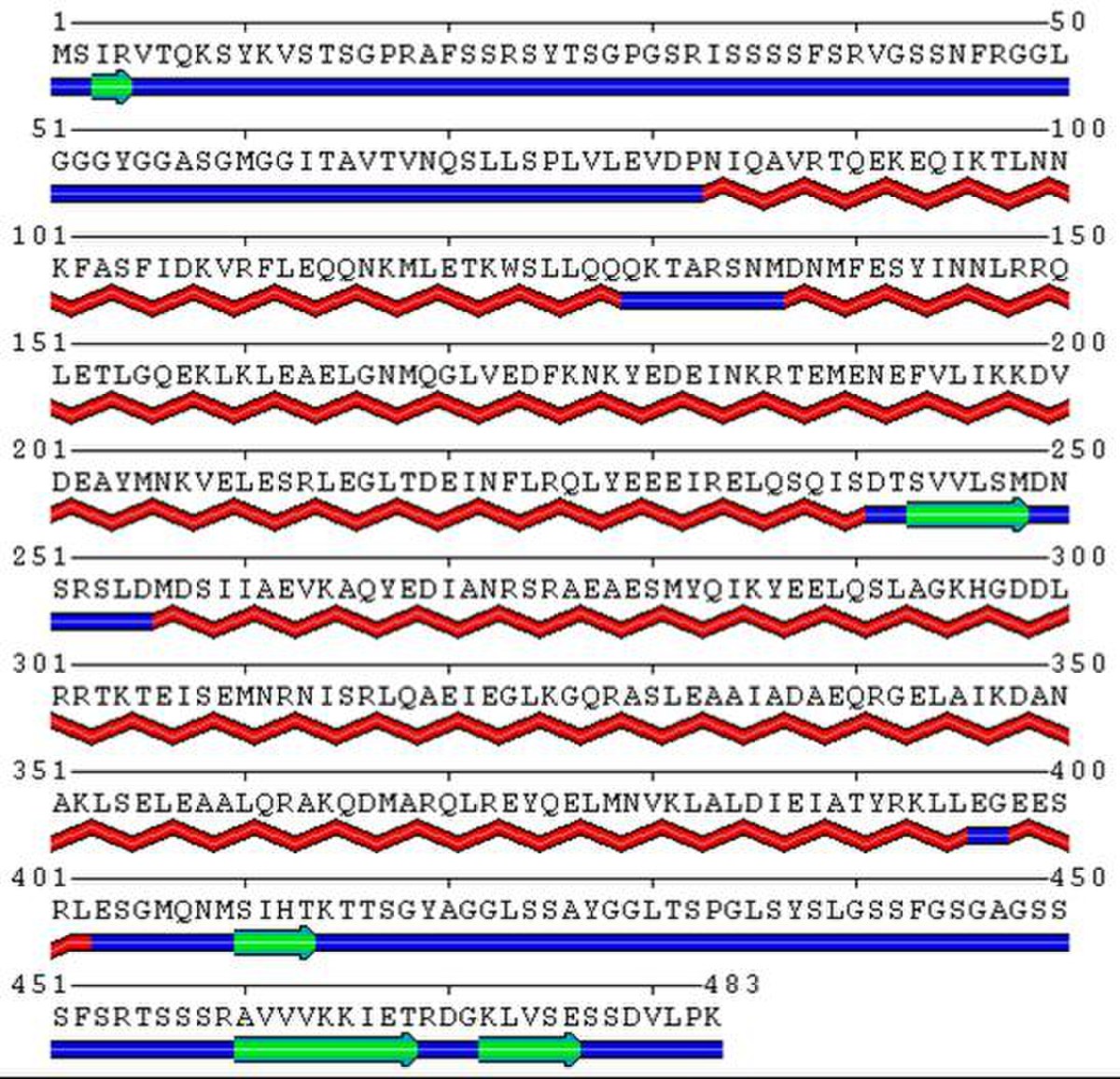
Du point de vue de sa structure, la kératine 8 est formée d’une portion centrale en hélice α, composée de quatre sous-domaines (1A, 1B, 2A et 2B) reliés entre eux par les portions (L1, L12 et L2). Pour s’agencer en dimère, la kératine 8 possède une forte homologie quant à la disposition de ces hélices α avec la kératine 18.

















































