Fossile - Définition
La liste des auteurs de cet article est disponible ici.
Localisation des sites fossilifères
Certaines régions du globe sont particulièrement connues pour l'abondance de leurs fossiles. Ces sites fossilifères d'une qualité exceptionnelle portent le nom de Lagerstätten (littéralement lieu de repos ou d'emmagasinage, en allemand). Ces formations résultent probablement de l'enfouissement de carcasses dans un environnement anoxique avec très peu de bactéries, ce qui a ralenti le processus de décomposition. Sur l'échelle des temps géologiques, les lagerstätten s'étendent du Cambrien à nos jours.
Parmi ces sites, on trouve notamment les marnes jurassiques de la Voulte-sur-Rhône (conservation des parties molles de céphalopodes en trois dimensions), les schistes de Maotianshan en Chine et ceux de Burgess en Colombie-Britannique, le calcaire lithographique de Solnhofen en Bavière. Celui-ci détient, par exemple, un des magnifiques exemples d'Archéoptéryx. Ces gisements fossilifères sont tellement rares qu'à chacune de leur découverte ou redécouverte, ils ont bouleversé la vision de la progression de la vie.
La fossilisation
La fossilisation des êtres vivants est en général un processus de minéralisation (remplacement des tissus vivants par des substances minérales) dans de la roche sédimentaire qui est la roche par excellence pour la conservation de fossiles. Dans des cas plutôt rares, on peut avoir une conservation de la matière organique (mammouth dans le pergélisol, momification dans du bitume, la diatomite (roche siliceuse), inclusion dans de l'ambre). Dans d'autres cas, ce ne sont que des traces d'activité biologique qui sont conservées (paléoichnologie).
Processus de décomposition
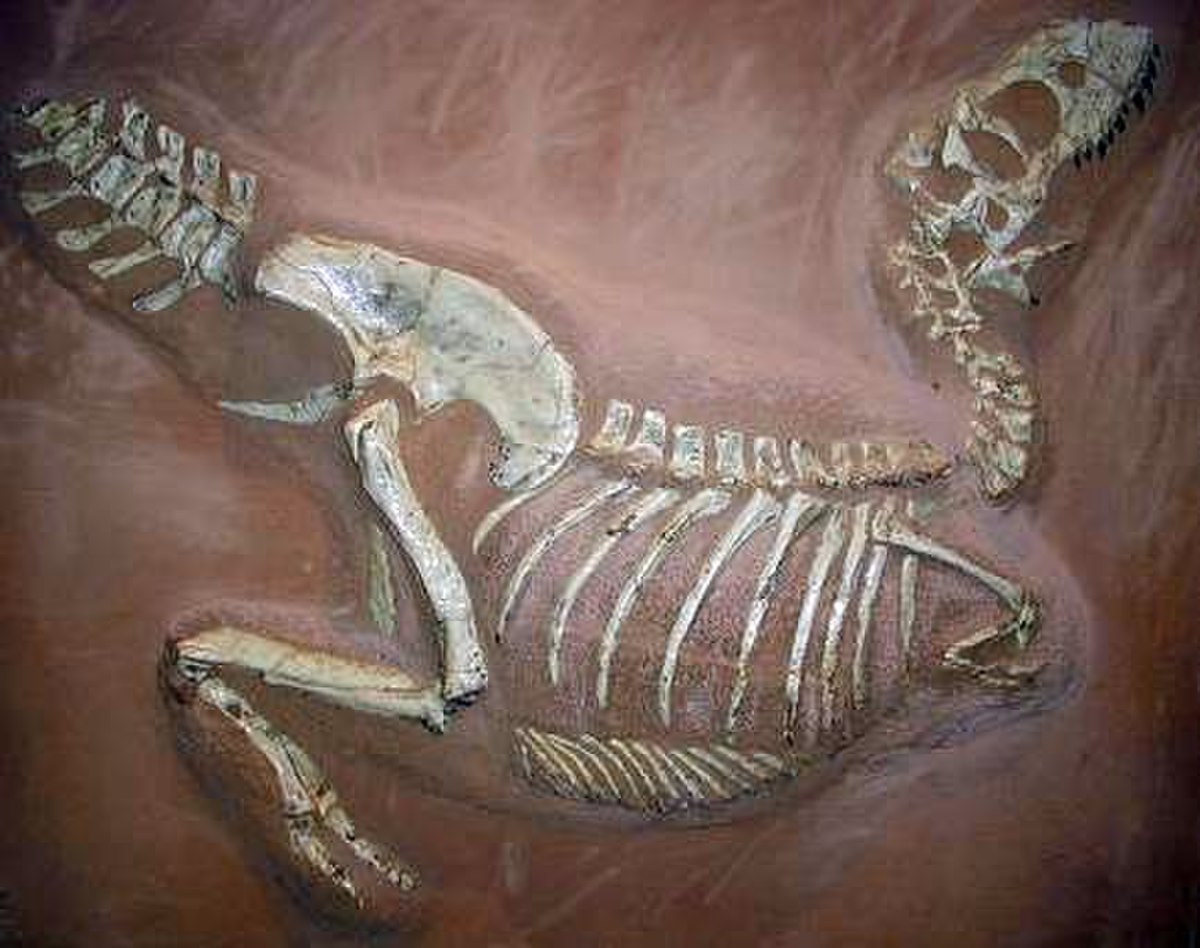
La capacité de conservation des fossiles est en grande partie due au processus de décomposition des organismes. Celui-ci explique pourquoi il est rare de retrouver des fossiles des parties molles organiques (60% des individus d'une communauté marine sont uniquement composés de parties molles). La présence des parties molles est alors le résultat de conditions sédimentologiques et diagénétiques exceptionnelles.
La biodégradation s'effectue de manière rapide et efficace grâce à une décomposition aérobie. Ainsi, il est nécessaire d'avoir un environnement anoxique pour pouvoir préserver des organismes légèrement minéralisés et composés de parties molles. Ceci exige une concentration élevée d'oxygène (106 moles pour 1 mole de carbone organique) :

La décomposition est la source principale de perte de données dans le registre fossile et la minéralisation est le seul moyen de la freiner. Les tissus peuvent se conserver sous la forme de perminéralisations (déchets organiques altérés) ou, quand la détérioration est prolongée, sous la forme d'empreintes. Si la décomposition est plus importante que la minéralisation, les tissus sont détruits et seuls les matériaux réfractaires (chitine, lignine ou cellulose) sont conservés.
La décomposition dans le registre fossile se caractérise à trois niveaux. Dans un premier temps, il s'agit d'identifier la décomposition et la perte d'information sur la structure de l'organisme fossilisé. Ensuite, il faut reconnaître les minéraux particuliers et les marqueurs géochimiques associés aux régimes particuliers de décomposition. Enfin, il faut garantir la préservation des microbes fossiles impliqués dans le processus.
La majeure partie de la matière organique se recycle pour la majeure partie à l'intérieur de la colonne d'eau, en particulier dans la zone euphotique. Une petite proportion de la matière organique produite sert à la formation des sédiments adjacents et est affectée par les modifications du flux organique (biostratinomique) telles que la photo-oxydation, l'activité microbienne et les organismes détritivores.
Processus de diagenèse fossile

En plus des lipides, la matière organique comprend également des biopolymères, comme les glucides, les protéines, la lignine et la chitine, dont certains seront utilisés pour sa consommation ou modifié par les organismes benthiques et les micro-organismes. Ceux qui ne sont pas utilisés pourront subir une polycondensation qui conduira à la formation de géopolymères qui s'intégreront au proto-kérogène (précurseur du kérogène). Lors de l'enfouissement des sédiments, la condensation s'accroît et l'insolubilité produit la lente conversion diagénétique du kérogène, constituant principal de la matière organique dans les sédiments anciens.
On trouve de grandes quantités de molécules organiques dans les sédiments et les roches sédimentaires. On les qualifie de marqueurs biologiques ou de « biomarqueurs ». Leur étude et leur identification nécessitent des techniques avancées d'investigation et d'analyse. Ces marqueurs conservent un registre très détaillé de l'activité biologique passée et ils sont liés aux molécules organiques actuelles. On trouve autant de sources possibles de marqueurs biologiques dans les échantillons que de molécules dans un organisme.
Une roche mère est un volume rocheux qui a généré et expulsé des hydrocarbures en quantité suffisante pour former une accumulation de pétrole et de gaz. La plupart des roches mères potentielles contiennent entre 0,8 et 2% de carbone organique. Il est couramment admis, comme limite basse, un pourcentage de 0,4% en volume de carbone organique pour la production d'hydrocarbures. Toutefois, la génération est plus efficace avec un pourcentage supérieur à 5-10%. La nature des hydrocarbures générés dépend essentiellement de la composition du kérogène, qui peut être composé de deux types de matières organiques : les débris de plantes terrestres - les sédiments libèrent alors du gaz - ou d'organismes aquatiques, comme les algues, le phytoplancton, le zooplancton - auquel cas ils forment alors du pétrole (si la maturation est suffisante).
Processus de destruction physico-chimique

La durabilité des squelettes dépend de leur résistance à la rupture et à la destruction par des agents chimiques, physiques et biotiques. Ces processus destructeurs peuvent être divisées en cinq catégories qui suivent plus ou moins l'ordre séquentiel : la désarticulation, la fragmentation, l'abrasion, la bioérosion et la corrosion/dissolution.
La désarticulation correspond à la désintégration de squelettes composés de plusieurs éléments le long des jointures ou des articulations préexistantes. Ce phénomène peut également se produire avant même la mort, comme la mue ou l'exuvie chez les arthropodes). Cette décomposition détruit les ligaments reliant les ossicules d'échinodermes en quelques heures ou quelques jours après la mort. Les ligaments, comme ceux des moules, composés de conchyoline, sont plus résistantes et peuvent rester intacts pendant des mois, en dépit de la fragmentation de la coquille.
La fragmentation se produit lors d'un impact par des objets physiques et par des agents biotiques, tels que les prédateurs ou les nécrophages. Certaines formes de rupture permettent d'identifier le prédateur. Les coquilles ont tendance à se briser le long de lignes de faiblesse pré-existantes, telles que les lignes de croissance ou d'ornementation. La résistance à la fragmentation dépend de plusieurs facteurs : la morphologie du squelette, la composition et la microstructure (notamment épaisseur et pourcentage de matière organique).
L'abrasion est le résultat du polissage et du concassage des éléments du squelette, qui produit un arrondissement et une perte des détails de la surface. Il y a eu des études semi-quantitatives sur les proportions de l'abrasion, en introduisant des coquilles dans un tambour rotatif, rempli de gravier siliceux. Le degré d'intensité est liée à plusieurs facteurs : l'énergie du milieu, le temps d'exposition, la taille de la particule abrasive et la microstructure du squelette.

La bioérosion ne peut se produire que si elle est associée à des fossiles reconnaissables, tels que les éponges Cliona ou les algues endolithiques. Son action destructrice est très importante dans les milieux marins peu profonds, où on peut observer une perte de masse allant de 16 à 20% dans les coquilles des mollusques actuels. Aucune étude ne montre toutefois si les proportions étaient les mêmes au Paléozoïque, quand les éponges cliona étaient moins abondantes.
La corrosion et la dissolution est le résultat de l'instabilité chimique des minéraux qui se trouvent dans la colonne d'eau et dans les pores des sédiments. La dissolution commence à l'interface sédiment-eau avant de continuer vers l'intérieur du sédiment. La bioturbation des sédiments favorise normalement la dissolution grâce à l'introduction d'eau de mer à l'intérieur du sédiment, ce qui permet également l'oxydation des sulfures.
Dans la pratique, il est difficile de distinguer les effets de l'abrasion mécanique, de la bioérosion et de la corrosion. Certains auteurs ont ainsi proposer le terme de corrasion pour indiquer l'état général des coquilles, comme le résultat d'une combinaison de ces processus. Le grade de corrasion est proportionnel à un indice général du temps durant lequel les restes ont été exposés à ces trois processus.
Diagenèse fossile
La compréhension des processus diagénétiques est essentielle pour l'interprétation correcte de la minéralogie originale, de la structure des squelettes et des coquilles, de leurs affinités taxonomiques et de la paléoécologie. L'un des problèmes auxquels nous sommes confrontés est très souvent de déduire ce qu'a été la minéralogie originale de groupes disparus (coraux bruts, archéocyathes, stromatopores...). La transition vers un état de fossile dépend surtout de la composition du squelette.
Nodule de carbonate et de calcaire lithographique

La préservation des parties molles est souvent associée à la précipitation des carbonates sous la forme de nodules stratifiés, comme pour le calcaire lithographique. Les nodules de carbonates sont composés de calcite ou de sidérite, et associés aux sédiments argileux riches en micro-organismes. Ils contiennent souvent des fossiles conservés dans leurs trois dimensions, et contiennent parfois même les restes fossilisés des parties molles. Leur taille varie entre 10 et 30 centimètres, même si certains atteignant les 10 mètres ont été retrouvés (dont un Plésiosaure complet). Le contenu de micro-organismes et leur décomposition sont les principaux facteurs qui contrôlent le degré d'anoxie, le potentiel d'oxydo-réduction et le pH. En présence d'oxygène, la respiration microbienne produit du CO2 qui s'accumule dans l'eau interstitielle des sédiments, favorisant la dissolution des carbonates :

En l'absence d'oxygène, les bactéries utilisent une série d'oxydants alternatifs dans le processus de la respiration (Mn, NO3-, Fe ou SO42-). Une fois que tous les oxydants ont disparu, la fermentation devient la réaction dominante et la production de méthane augmente. Le calcaire lithographique se forme dans un environnement marin ou lacustre et présente sous forme de fines bandes à grain fin. On peut citer comme exemple, le célèbre calcaire de Solnhofen datant du Jurassique et contenant des fossiles d'Archaeopteryx. Les dépôts de carbonate peuvent provenir de sources biogéniques (comme les algues calcaires) ou d'un précipité chimique.
Squelette d'aragonite
Normalement, l'aragonite se transforme en calcite à travers un processus de dissolution ou de calcification. Si les eaux du gué ne sont pas saturés en carbonates, il se produit une dissolution totale du squelette et des chairs par la calcite. L'espace vide reproduit le moule d'une coquille vide et la structure de cette dernière n'est pas conservée. Il peut se former des druses avec des cristaux dirigés vers le centre. La durée de ce processus est variable. Dans le cas de la calcification, le squelette des coquilles conserve son ancienne structure (en couches ou lamelles). Il se peut même que soient préservés les cristaux d'aragonite, ce qui nous donne des renseignements très utiles. Ce remplacement se fait progressivement et respecte la structure d'origine.
Squelette de calcite

En général, les squelettes fossiles qui étaient constitués de calcite, conservent souvent leur composition originale (à moins qu'ils ne se silicifient ou ne se dolomitisent). La teneur en magnésium a tendance à diminuer, de sorte qu'il puisse y avoir une altération diagénique, soit à forte, soit à faible teneur en calcite. Il existe des techniques spéciales, telles que la cathodoluminescence, pour déterminer son contenu original à partir des zones qui ont conservé leur composition originale.
Squelette calcaire
Les squelettes de carbonate de calcium peuvent se transformer en apatite sans modification de la morphologie externe. Dans les milieux naturels, cette modification diagénique est associée à des dépôts de phosphate. La transformation bactérienne des organismes calcaires en apatite a été démontrée en laboratoire. Ces observations et ces expériences suggèrent, dans un premier temps, que le phosphore nécessaire pour remplacer le carbonate par de l'apatite provient des micro-organismes des sédiments. Par ailleurs, il semble que les micro-organismes (bactéries, algues, champignons) favorisent la décomposition, en libérant des ions phosphates et en acidifiant l'eau interstitielle des sédiments. Cette acidification, qui peut être très localisée, favorise la dissolution des carbonates. Le phosphate libéré se combine avec le calcium pour former de l'apatite, préférentiellement à l'interface entre le carbone et le micro-organisme remplaçant le carbonate dissous. Ce remplacement préserve l'apparence originale de la coquille et le fluor joue un rôle important en ce qui concerne la composition finale en carbonate-fluor-apatite.
Squelette de silice

Le phosphatisation de la silice primaire apparaît aussi sur certains squelettes de radiolaires, bien que ce processus ne soit pas encore bien connu à l'heure actuelle. L'examen microscopique d'échantillons de phosphorites montre que de nombreux micro-organismes sans carapace minérale (algues, champignons, bactéries) se minéralisent comme l'apatite, bien qu'ils n'aient aucun précurseur minéral. Un exemple bien connu est le coprolithe phosphaté, où la matière organique est elle-même remplacée par de l'apatite qui conserve la forme exacte de l'objet. La phosphatisation des parties molles est également fréquente, notamment chez de nombreux arthropodes (copépodes, ostracodes) où des nodules calcaires et phosphatés apparaissent au sein de calcaire nodulaire ou de coprolithes de grands vertébrés.
Des études sur les phosphorites et sur la synthèse expérimentale de l'apatite ont abouti à une estimation des conditions probables de fossilisation de l'apatite. En raison de son besoin de stabilité, l'apatite se forme de préférence dans un environnement déficient en oxygène, parfois même dans des conditions totalement réductrices, comme l'indique la présence fréquente de pyrite à proximité. Cet environnement est atteint facilement dans les milieux où l'on trouve beaucoup de matière organique qui est la principale source de phosphore.
La silice peut remplacer la calcite et l'aragonite des coques et perminéraliser le bois. Il peut également se former des nodules et des couches de silex, en remplaçant les sédiments carbonés, en précipitant directement ou en remplissant les fossiles ou les inclusions. La coque peut alors être remplacées par une croûte blanche granuleuse, par une couche finement granuleuse ou par des anneaux concentriques de silice.
Fossile pyritisé

La pyrite sédimentaire est une composante mineure des sédiments clastiques marins. Les études actuelles sur les sédiments ont montré que la formation de la pyrite authigénique a lieu au tout début de la diagenèse, à quelques centimètres au-dessous de l'interface eau-sédiments. Une augmentation du nombre de micro-organismes et/ou de la profondeur d'enfouissement empêche la diffusion de l'oxygène dans les sédiments et les micro-organismes sont obligés de respirer en anaérobie. La minéralisation empêche la perte d'information relative à la décomposition de macro-organismes et la précipitation de la pyrite, au début de la diagenèse, est un moyen important pour la préservation des fossiles. Dans les tissus mous, comme les muscles et la chitine, il peut se produire un pyritisation au début de la diagenèse. Lorsque la décomposition est plus avancée (mais avant que ne se produise la formation de la pyrite), les tissus mous seront détruits et seuls les composés biologiques résistants (appelés réfractaires), comme la cellulose et la lignine, sont préservés. Les parties biogéniques dures, telles que les coquilles (composées de carbonate de calcium et de magnésium) et les os (phosphate de calcium) sont quelques-unes des structures biologiques les plus résistantes à la décomposition. Sur les deux, le carbonate de calcium est le plus instable et il est donc plus probable qu'il soit remplacé par la pyrite.
La formation de la pyrite est contrôlée par la concentration en carbone organique, en sulfate et en minéraux détritiques ferreux. Dans un environnement marin normal, les minéraux ferreux et les sulfates sont présents en abondance et la formation de pyrite est contrôlée par l'approvisionnement en carbone organique. Toutefois, dans les milieux en eau douce, la formation de pyrite est très limitée par la faible concentration en sulfates.
Plante fossile

Les différentes parties des plantes (branches, racines, feuilles, pollen, fruits, graines) se détachent pour certaines au cours de leur vie, et pour les autres après leur mort. Une bonne compréhension des processus de dispersion qui affectent la dispersion de ces parties est très important pour interpréter correctement les associations paléofloristiques. Les études sur la dispersion des feuilles par le vent montrent qu'elle dépend de leur poids et de leur forme. Les débris végétaux se conservent soit par préservation du matériel original, soit par carbonisation, soit par perminéralisation.
La perminéralisation se produit après l'enfouissement, quand les espaces vides situés à l'intérieur d'un organisme (espaces remplis de liquide ou de gaz, quand il est en vie) se remplissent d'eaux souterraines riches en minéraux et que ces minéraux précipitent en comblant les espaces vides. Ce processus ne peut se produire que dans de très petits espaces, notamment au sein de la paroi cellulaire d'une cellule végétale. Une perminéralisation à petite échelle peut produire des fossiles avec de très nombreux détails. Pour que la perminéralisation se produise, il faut que l'organisme soit recouvert par les sédiments peu de temps après la mort ou peu après le début du processus de décomposition. La vitesse de dégradation des restes, une fois recouverts, détermine les futurs détails du fossile. Ainsi, alors que certains fossiles ne comprennent que des ossements ou des dents, d'autres contiennent des traces fossiles de peau, de plumes ou même de tissus mous.

















































