Extracellular signal-regulated kinases - Définition
La liste des auteurs de cet article est disponible ici.
Fonctions
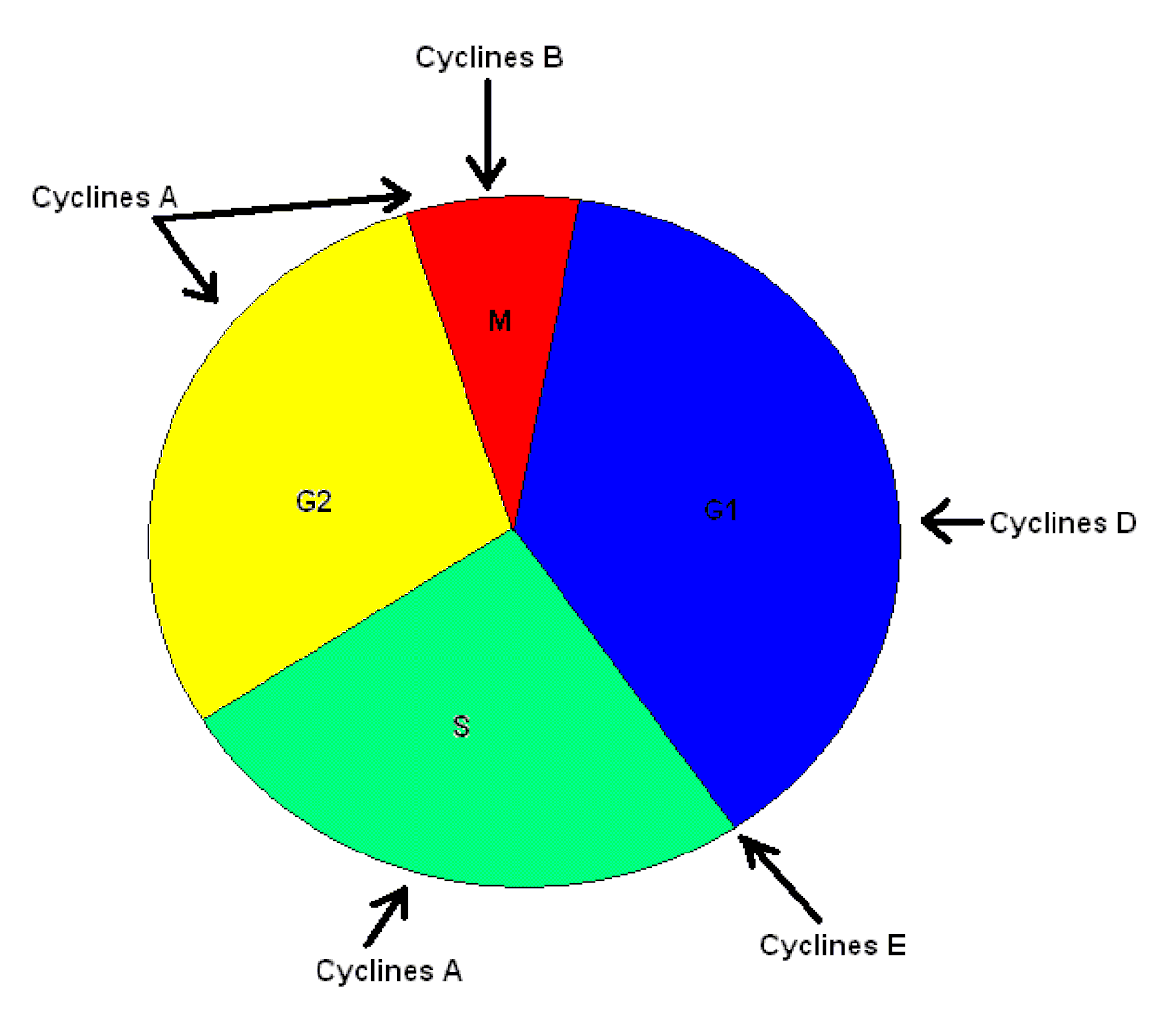
Les MAP kinases ont un rôle important à jouer dans la croissance, la prolifération et la différentiation cellulaires; elles sont également «des régulateurs ubiquitaires» (se retrouvent dans pratiquement tous les types de cellule de l’organisme). Le signal de transduction engendré par diverses molécules (facteurs mitogènes, stimuli de différentiation et cytokines) active en bout de ligne ERK1/2 qui phosphorylent plusieurs substrats cytoplasmiques et nucléaires nécessaires à la transcription de nombreux gènes afin de passer de la phase G1 à la phase S dans le processus de division cellulaire (Voir figure 2). On retrouve ERK1/2 chez plusieurs organismes, notamment chez les levures, chez les eucaryotes, chez les grenouilles et chez les mammifères où ces protéines ont été étudiées. Chez ces organismes, cette voie de signalisation, Ras-dependent extracellular signal-regulated kinase (ERK)1/2 mitogen-activated protein (MAP) kinase pathway, est évolutivement bien conservée et possède les mêmes fonctions. De plus, il est possible de remarquer que cette voie a été adaptée à différentes fins. Ainsi, les levures utilisent la voie de signalisation des MAP kinases ERK1/2 pour répondre lors de stimuli, comme les phéromones, pendant la période de reproduction sexuée. Une levure prolongera alors ses hyphes afin de rejoindre les hyphes de l’autre levure afin de se reproduire de façon sexuée. Il y a aussi les petites mouches à fruits (drosophiles) qui utilisent cette voie de transduction pour la différentiation des photorécepteurs de la rétine. Les plantes à fleurs, quant à elles, initient une défense contre les pathogènes grâce au signal reçu par cette voie de signalisation.
Identité des protéines ERK1/2 dans différentes banques de données
Protein: ERK-1: Rattus norvegicus: P21708
Human: P27361
ERK-2: Rattus norvegicus: NP_446294
Human: NP_002736
Gene: ERK-1: Rattus norvegicus: GeneID: 50689
Human: GeneID: 5595
ERK-2: Rattus norvegicus: GeneID: 116590
Human: GeneID: 5594
OMIM (Human): ERK-1: *601795
ERK-2: *176948
SwissProt (Human): ERK-1: P27361
ERK-2: P28482
Pathologies associées
ERK1/2 se retrouvant dans la plupart des cellules de l’organisme et étant des protéines très importantes dans la différentiation, la prolifération et la croissance cellulaires, une surexpression ou une absence de leurs niveaux cause de très graves conséquences. En premier lieu, il faut comprendre ce qui se passe dans une cellule normale afin d’envisager les effets d’un dérèglement cellulaire. Lorsqu’il y a une surexpression ou une suractivation de ERK1/2 dans la cellule, les inhibiteurs p15 et p16 augmentent dans la cellule et engendrent alors l’arrêt du cycle cellulaire. Ce précédé a été étudié dans certains types de cellules tels les fibroblastes, les cellules de Schwann, les hépatocytes, les lymphocytes T, les kératinocytes, les astrocytes et les cellules épithéliales de l’intestin. Donc, ces inhibiteurs forment le mécanisme de protection mis en place pour éviter la prolifération cellulaire anormale causée par un signal mitotique trop fort et ainsi permet d’éviter la formation de cancers. Un dérèglement de ce mécanisme et l’activation constitutive de ERK1/2 causeront possiblement le développement d’une tumeur.
En deuxième lieu, une diminution ou même carrément l’absence de ERK1/2 provoque également des effets néfastes. En effet, une mutation sur le gène de ERK1 ou sur le gène de ERK2 entraîne, par exemple, une baisse de la prolifération des lymphocytes T en présence d’un agent infectieux, car les lymphocytes T possède le complexe AP-1 qui sert à la production d’interleukine-2 essentiel à la croissance de ceux-ci. Alors, le système immunitaire n’arrive plus à combattre normalement contre l’infection. De plus, des études faites chez la souris dont on avait injecté la toxine létale de l’anthrax, ont montré une perturbation dans la voie des MAP kinase des macrophages. Une façon subtile d’attaquer le système immunitaire et de survivre dans l’organisme. Par ailleurs, une autre étude menée sur des souris knock-out Erk1-/-, ne présente aucune anomalie dans la différentiation thymique, mais chez d’autres types de cellules comme les kératinocytes, on remarque une baisse de prolifération en réponse aux agents mitotiques. De plus, une déficience de ERK-1 diminue la progression du papillome de stade 2 induit par des produits chimiques sur la peau de la souris. Par contre, lorsqu’une délétion est faite dans le gène ERK-2, alors on remarque un blocage dans la maturation des thymocytes. Ce qui fait croire que ERK-1 n’arrive pas à compenser pour le manque de ERK-2. Pour continuer, chez des souris ayant eu un infarctus, la perte de leur allèle Erk2 provoque une augmentation de la mort des cellules cardiaques. Par conséquent, ERK1/2 sont des protéines importantes dans la survie cellulaire.
En conclusion, les effets individuels de ERK-1 et ERK-2 ne sont pas encore bien connus à ce jour. Mais il est évident que ERK-2 possède des effets sur le développement embryonnaire, car, chez la souris, si la protéine ERK-2 n’est pas présente, il y aura létalité embryonnaire après 7,5 jours. Le souriceau ne viendra jamais à terme. Il n’est pas possible de vérifier ces résultats chez l’humain pour des questions d’éthique. Il est impossible de pouvoir analyser tous les embryons humains morts afin de suspecter un lien avec ERK1/2. Bref, ERK1/2 doivent être activées en quantités suffisantes pour avoir ses effets bénéfiques dans l’organisme.

















































