Extinction Crétacé-Tertiaire - Définition
La liste des auteurs de cet article est disponible ici.
Introduction


L’Extinction Crétacé-Tertiaire ou Extinction K/T qui marque la fin du Crétacé est une extinction massive et à grande échelle d'espèces animales et végétales qui s'est produite il y a 65,5 Ma environ, dans une courte période de temps à l'échelle géologique. Cet évènement marque la fin de l'ère mésozoïque et le début de l'ère cénozoïque. Largement connue comme l’extinction K-T, elle est associée à une signature géologique connue sous le nom de limite K-T, habituellement une couche mince d'argile présentant un taux anormal d'Iridium que l'on retrouve dans diverses régions du monde. K est l'abréviation traditionnelle pour la période du crétacé (dérivé du nom allemand Kreidezeit), et T est l'abréviation du tertiaire, terme historique qui désigne la période maintenant couverte par les périodes paléogène et néogène. L'emploi du terme « Tertiaire » étant maintenant déconseillé comme unité formelle de temps ou de roche par la Commission internationale sur la stratigraphie, l'évènement de K-T est maintenant désigné comme l'extinction du Cretacé-Paléogène (ou K-Pg) par beaucoup de chercheurs.
La plupart des fossiles de dinosaures non-aviaires se trouvent au-dessous de la limite du K-T, ce qui indique que les dinosaures non-aviaires se sont éteints juste avant, ou pendant l'évènement. On a découvert quelques fossiles de dinosaures au-dessus de la limite K-T, mais l'on pense que cela est dû à un remaniement des sédiments, c'est-à-dire que l'érosion a ramené les fossiles à la surface puis ils ont été recouvert par un dépôt de sédiments plus récents. Les mosasaures, les plésiosaures, les ptérosaures et de nombreuses espèces de plantes et d'invertébrés se sont également éteints. Les clades de mammifères et d'oiseaux ont survécu avec peu d'extinctions, et une radiation évolutive des taxons du Maastrichtien s'est produite bien après la limite. Les taux d'extinctions et de radiations ont varié à travers différents clades.
Les théories scientifiques expliquent les extinctions de K-T par un ou plusieurs évènements catastrophiques, tels que des impacts massifs d'astéroïdes , ou une activité volcanique accrue. La datation de plusieurs cratères d'impact (comme l'impact de Chicxulub) et d'une activité volcanique massive (dans les trapps du Deccan), coïncide avec la période approximative de l'évènement d'extinction. Ces évènements géologiques auraient réduit la lumière du soleil et gêné la photosynthèse, menant à une rupture massive dans l'écologie de la terre. D'autres chercheurs croient que l'extinction a été plus progressive, résultant de changements plus lents du niveau de la mer ou de climat.
En mars 2010, un groupe de 41 scientifiques se sont accordés dans la revue Science sur le fait que la chute de l'astéroïde à l'origine du cratère de Chicxulub avait été l'événement déclencheur de l'extinction K-Pg.
Schémas d'extinction
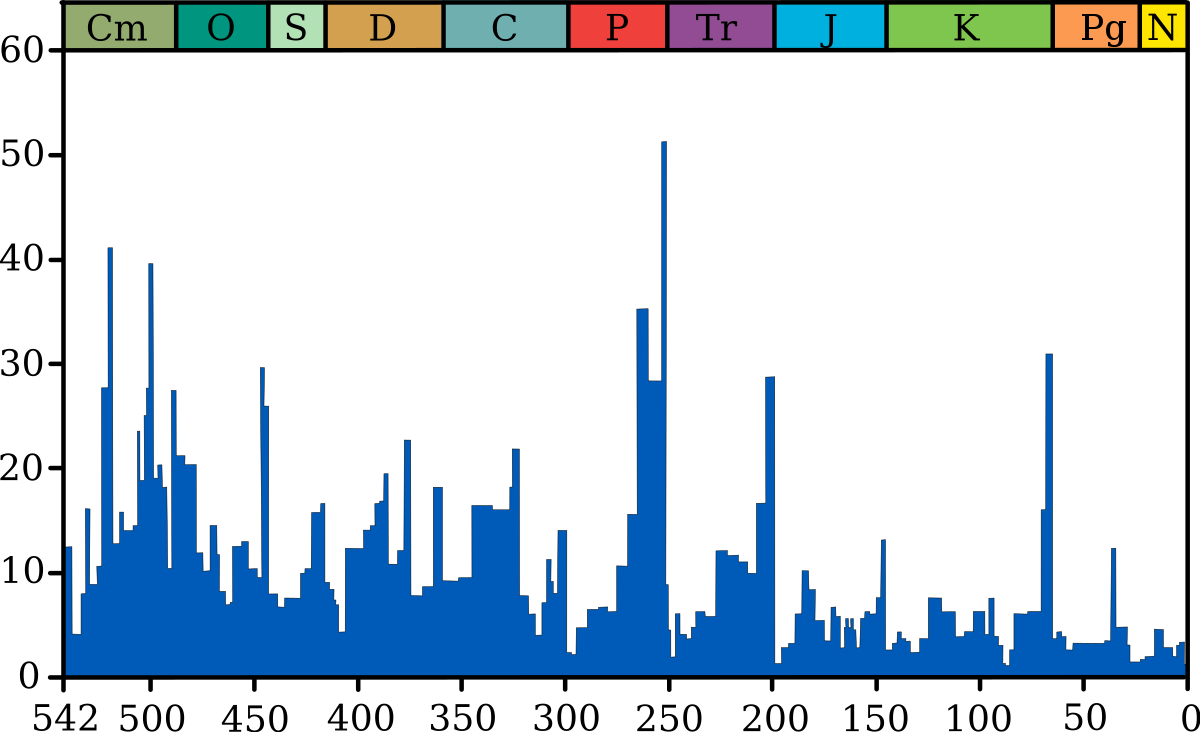
Millions d'années O-S Late D P-Tr Tr-J K-T |
Le graphique bleu indique le pourcentage apparent (pas en nombre absolu) de genres d'animaux marins ayant disparus au cours d'un intervalle de temps. Il ne représente pas toutes espèces marines, mais seulement les espèces marines fossiles. Les 5 plus grandes extinctions sont liées, voir les extinction massives pour plus de détails.
Source et information sur le graphique
Bien que l'évènement de la limite K-T ait été de grande ampleur, il y eut une variabilité significative du taux d'extinction entre les différents groupes d'espèces. On suppose que des particules atmosphériques ont bloqué la lumière du soleil, réduisant la quantité d'énergie solaire pouvant atteindre la terre. Les effectifs des espèces dépendant de la photosynthèse ont donc diminué, certaines espèces se sont même éteintes. Vers la fin du crétacé, les organismes qui photosynthétisent, dont le phytoplancton et les plantes terrestres, étaient à la base de la chaîne alimentaire comme c'est le cas aujourd'hui. L'extinction d'espèces végétales alors dominantes a causé un remaniement important de ce groupe d'organismes. L'observation suggère que les animaux herbivores se sont éteints quand les plantes dont ils dépendaient pour se nourrir sont devenues rares ; en conséquence, les prédateurs supérieurs comme le Tyrannosaure ont également péri.
Les coccolithophorides (des algues unicellulaires microscopiques) et les mollusques, incluant les ammonites, les rudistes, les escargots d'eau douce et les moules, ainsi que les organismes dont la chaîne alimentaire inclut ces animaux à coquilles se sont éteints ou ont connu de lourdes pertes. Par exemple, on pense que les ammonites étaient la nourriture principale de mosasaures, un groupe de reptiles marins géants qui se sont éteints précisément à cette période.
Les omnivores, les insectivores et les charognards ont survécu à l'évènement d'extinction, peut-être en raison de la plus grande disponibilité de leurs sources de nourriture. À la fin du crétacé il ne semble y avoir eu aucun mammifère purement herbivore ou purement carnivore. Les mammifères et les oiseaux qui ont survécu à l’extinction se sont nourris d'insectes, de vers, et d'escargots, qui eux-mêmes se nourrissaient de matière morte végétale ou animale. Les scientifiques pensent que ces organismes ont survécu à l'effondrement des chaînes alimentaires basées sur les plantes parce qu'ils se sont nourris de détritus ou d'autres matériaux organiques non vivants.
Dans les biocénoses des cours d'eau, peu de groupes d'animaux se sont éteints parce que ces communautés dépendent moins directement des plantes vivantes pour leur nourriture et plus des détritus qui ruissellent de la terre, ce qui les protégea de l'extinction. Des mécanismes semblables, mais plus complexes ont été observés dans les océans. Les extinctions ont été plus nombreuses parmi des animaux vivant dans la zone pélagique, que parmi des animaux des fonds sous-marins. Les animaux de la zone pélagique dépendent presque entièrement de la production primaire du phytoplancton vivant, alors que des animaux vivants dans les fonds marins s'alimentent de détritus ou peuvent passer à une alimentation constituée uniquement de détritus.
Les plus grands survivants aérobies de l'évènement, les crocodiliens et les champsosaures, étaient semi-aquatiques et ont eu accès aux détritus. Les crocodiliens modernes peuvent vivre comme charognards et survivre pendant des mois sans nourriture. En outre, dans leur jeunesse, ils se développent lentement et se nourrissent pendant leurs premières années en grande partie d'invertébrés et d'organismes morts ou de fragments d'organismes. Ces caractéristiques ont permis la survie de ces espèces à la fin du Crétacé.
Après l'évènement du K-T, la biodiversité a eu besoin d'un temps substantiel pour récupérer, en dépit de l'existence de nombreuses niches écologiques.
Microbiota
La limite K-T présente un des taux de disparitions et d'apparitions d'espèces les plus spectaculaires parmi celles enregistrées sur des espèces de fossiles connus. Ce phénomène a d'ailleurs particulièrement touché les nanoplanctons à l'origine des dépôts calcaires du Crétacé. Et de fait, de très nombreuses espèces, particulièrement parmi celles qui ont disparu, ont été remplacées par l'apparition de nouvelles. L'analyse statistique des disparitions des espèces marines suggère que la diminution de la diversité a davantage été provoquée par une forte hausse des extinctions plutôt que par une diminution de l'apparition de nouvelles espèces, par un mécanisme naturel et permanent appelé spéciation.
Les données concernant les phytoplanctons appelés dinophytes ne sont pas aussi bien expliquées durant cette période de transition du Crétacé au Paléogène, principalement parce que seuls les kystes fournissent de bonnes traces fossiles dans ce groupe, or les espèces de dinophytes ne passent pas toutes par le stade de kyste, ce qui est probablement à l'origine d'une sous-estimation de leur diversité. Les études récentes tendent à indiquer qu'il n'y aurait pas eu de variations significatives des dinophytes au niveau des couches de dépôts fossiles qui constituent la limite entre Crétacé et Tertiaire.
Des traces géologiques de radiolaires existent depuis au moins l’époque ordovicienne, et leurs squelettes siliceux fossiles peuvent être facilement repérés et suivis au cours de la limite K-T. Il n'y a aucune preuve d’extinction de masse de ces organismes, et en raison du refroidissement des températures au début du paléocène, on note une productivité élevée pour ces espèces au niveau des latitudes élevées de l'hémisphère Sud. En ce qui concerne les diatomées, environ 46% des espèces ont survécu à la transition crétacé-paléocène, ce qui suggère un taux de renouvellement significatif des espèces, mais pas une extinction catastrophique au passage de la limite K-T.
La présence des foraminifères planctoniques au passage de la limite K-T a été étudiée dès les années 1930. Ces recherches, stimulées par la possibilité d'un d'impact cosmique, produisirent de nombreuses publications détaillant leur extinction au cours de cette période de transition. Cependant, des discussions sont en cours entre ceux qui croient que les données fossiles indiquent une extinction substantielle, et ceux qui pensent que les données fossiles montrent de multiples extinctions et apparitions d'espèces au passage de cette limite. Parmi ces espèces, les espèces benthiques c'est-à-dire celle de haute mer, semblent toutes s'éteindre. On pense que la biomasse des océans a diminué après les extinctions qui ont eu lieu sur les côtes et que ces foraminifères dépendaient des débris organiques pour leur alimentation. Lorsque plus tard le nombre d'espèces de microorganismes marins a réaugmenté, on observe une augmentation concomitante du nombre d'espèces de foraminifères benthiques, vraisemblablement en raison de l'augmentation des ressources alimentaires. Autrement dit, le rétablissement des populations de phytoplancton au début du paléocène a fourni la source de nourriture nécessaire pour alimenter de nouveau de grandes populations de foraminifères benthiques, qui se nourrissent toujours principalement de détritus. Au début du paléocène, le rétablissement final de ces populations benthiques s'est réalisé en plusieurs étapes qui ont duré plusieurs centaines de milliers d'années.
Invertébrés marins

Il y a une grande variabilité en ce qui concerne les taux d'extinction des invertébrés marins au passage de la limite K-T. Le nombre de fossiles et de sites fossilifères connus est faible, de ce fait les taux d'extinction calculés à partir des données récoltées peuvent ne pas correspondre à la réalité, les surestimant.
Les ostracodes, une classe de petits crustacés qui étaient très communs lors du Maastrichtien supérieur, ont laissé des traces fossiles dans un grand nombre d'endroits. Un examen de ces fossiles prouve que la diversité des ostracodes est plus faible au cours du paléocène qu'à n'importe quelle autre période du Tertiaire. Cependant, dans l'état actuel de la recherche, on ne peut pas déterminer si ces extinctions se sont produites avant ou pendant l'intervalle de la limite.
Chez les coraux du Crétacé tardif, environ 60 % des genres appartenant à l'ordre des Scleractinia (coraux durs) n'ont pas réussi à passer la limite K-T et à atteindre le paléocène. L'analyse approfondie des extinctions de coraux de cette époque montre qu'environ 98 % des espèces coloniales qui habitaient les eaux tropicales chaudes et peu profondes se sont éteintes. Les coraux solitaires, qui ne forment généralement pas de récifs et habitent des régions plus froides et plus profondes de l'océan (au-dessous de la zone photique) ont moins été affectés par la limite K-T. Les espèces des colonies coralliennes dépendent d'une symbiose avec des algues photosynthétiques, dont les populations se sont effondrées en raison des évènements entourant la limite K-T. Cependant, l'explication de l'extinction K-T et du rétablissement au paléocène par l'utilisation des données des fossiles de coraux doit être relativisée en raison des changements qui se sont produits dans les écosystèmes de coraux au passage de la limite K-T.
Les nombres de genres de céphalopodes, d’échinodermes et de bivalves ont diminué significativement après la limite K-T. La plupart des espèces de brachiopodes, un petit phylum d'invertébrés marins, ont survécu à l'évènement K-T et se sont diversifiés au début du paléocène.

Mises à part la sous-classe des Nautiloidea, représentée aujourd'hui par les espèce des nautiles et un groupe issu d'une évolution de ceux-ci, à savoir les Coleoidea qui regroupe entre autres les octopodes, calmars, et seiches, toutes les autres espèces de mollusques de la classe des céphalopodes se sont éteintes à la limite K-T. Parmi celles-ci, il y avait les espèces du super-ordre Belemnoidea et les ammonites, une sous-classe de céphalopodes à coquille univalve, très diversifiés, dont les spécimens était nombreux, et à la répartition large. Des études scientifiques réalisée sur le sujet ont précisé que la stratégie reproductrice des nautiloïdes survivants, qui s'appuyait sur des œufs moins nombreux et plus gros, a joué un rôle dans leur conservation par rapport aux ammonites au cours de l'extinction. Les ammonites, elles, utilisaient d'une stratégie planctonique, c'est-à-dire que les œufs et et larves étaient aussi nombreuses que petites, ce qui aurait été très défavorable. Des chercheurs ont montré que suite de la disparition des ammonites, les nautiloïdes ont connu une radiation évolutive avec des formes et des complexités de coquille qui n'avaient été auparavant observés que chez les ammonites.
Environ 35 % des genres d'échinodermes se sont éteints à la limite K-T, ce qui est peu car ce sont les taxons qui prospéraient à la fin du crétacé dans les eaux peu profondes, à faible latitude, qui ont eu le taux d'extinction le plus élevé. Au niveau des latitudes moyennes, les échinodermes d'eaux profondes ont été beaucoup moins affectés par l'extinction. Ce serait une perte d'habitat qui serait responsable de ces extinctions, spécifiquement la submersion des récifs d'eau peu profonde existant à ce moment-là, lors d'un épisode transgressif survenu à une période proche de celle de l'évènement d'extinction.
D'autres groupes invertébrés marins, notamment des bivalves comme les rudistes et les inocérames, se sont également éteints à la limite K-T.
Les vertébrés marins
Un nombre substantiel de fossiles de poissons ont été découverts. Cela fournit de bonnes bases pour comprendre les modèles d'extinction de ces classes de vertébrés marins. Parmi les Chondrichthyes, approximativement 80 % des familles de requins, de raies et autres poissons cartilagineux ont survécu à l'évènement d'extinction. Par ailleurs moins de 10% des familles de poissons osseux, c'est-à-dire les téléostéens, se sont éteintes. Mais, près de l'Antarctique, Sur l'île de Seymour (en), un site fossile daté de la période précédant immédiatement l'évènement présente des preuves d'une mort massive chez des poissons osseux. On spécule que les poissons ont subi un stress environnemental avant l'évènement de la limite K-T et que ce dernier a dû précipiter l'extinction de masse. Cependant, il semble que les environnements marins et d'eau douce ont atténué l'effet de l'extinction sur les poissons.
Invertébrés terrestres
Les dommages causés par les insectes sur les feuilles fossilisées de plantes à fleurs de quatorze emplacements en Amérique du Nord ont été employés comme indicateur de la diversité des insectes à travers la limite K-T et analysés pour déterminer le taux d'extinction. Les chercheurs ont constaté que les sites du crétacé, avant l'évènement d'extinction, avaient une grande richesse en plantes et diverses formes d'alimentation par les insectes. Cependant, au début du paléocène, la flore étaient relativement diverse avec peu de prédation des insectes, même 1,7 million d'années après le phénomène d'extinction.
Plantes terrestres
Il existe des preuves dominantes d'une rupture globale des groupes de plantes à la limite K-T. Cependant, il y a eu d'importantes disparités selon les régions observées dans la succession des plantes. En Amérique du Nord, les données suggèrent une dévastation massive des plantes dans les sections de limite K-T, bien qu'il y ait aussi eu des changements mégafloraux ⇔ substantiels avant la limite.
Dans les latitudes élevées de l'hémisphère sud, comme la Nouvelle-Zélande et l'Antarctique, la décroissance de masse de la flore n'a causé aucun renouvellement significatif dans les espèces, mais des changements dramatiques à court terme dans l'abondance relative des groupes de plantes. En Amérique du Nord, approximativement 57 % des plantes se sont éteintes. La redressement des plantes au paléocène a commencé par des re-colonisations par des espèces de fougères, qui se présentent sous forme de pointe de fougère (en) au niveau géologique ; on a observé ce même type de re-colonisation de fougères après l'éruption du mont Saint Helens en 1980.
En raison de la destruction en masse des plantes à la limite K-T, il y a eu prolifération des organismes détritivores comme les mycètes qui n'ont pas besoin de photosynthèse et utilisent les nutriments de la végétation en décomposition. La domination des espèces fongiques a duré seulement quelques années tandis que l'atmosphère se dégageait et qu'il y avait abondance de matière organique. Une fois l'atmosphère dégagée, les organismes photosynthétiques comme les fougères et d'autres plantes sont revenues. La polyploïdie semble avoir augmenté la capacité des plantes à fleur à survivre à l'extinction, probablement parce que les copies additionnelles du génome que ces plantes possédaient leur ont permis de s'adapter plus facilement aux conditions environnementales en pleine mutation qui ont suivi l'impact.
Amphibiens
Il n'y a aucune trace d'extinctions de masse d'amphibiens à la limite K-T, et il y a une preuve irréfutable que la plupart des amphibiens ont survécu à l'évènement relativement indemnes. Plusieurs études approfondies des genres de salamandres dans les lits fossiles du Montana montrent que sur sept genres, six étaient inchangés après l'évènement.
Les espèces de grenouilles semblent avoir survécu jusqu'au paléocène avec peu d'extinction d'espèces. Cependant, les fossiles pour des familles et des genres de grenouille sont irréguliers. Une étude approfondie de trois genres des grenouilles du Montana montrent qu'elles n'étaient pas changées par l'événement de K-T et qu'elles ont été survécu apparemment sans changement. Les données montrent peu ou pas d'évidence d'extinction de familles amphibies qui encadrent l'événement de K-T. La survie amphibie a résulté de la capacité du clade à se chercher un l'abri dans l'eau ou à construire des terriers en sédiments, sol, bois, ou sous des roches.
Reptiles non-archosaures

Les deux taxa de reptiles non-archosaures vivants, les testudines (tortues) et lépidosauriens (serpents, lézards, et lézard verts), ainsi que les choristodères (des archosauromorphes semi-aquatiques qui se sont éteints au début du miocène) ont survécu au passage de la limite K-T. Plus de 80% des espèces de tortues du crétacé ont traversé la limite K-T. De plus, les six familles de tortues qui existaient à la fin du crétacé ont survécu au tertiaire et sont représentées par des espèces actuelles.
Les lépidosauriens vivants incluent les rhynchocéphales et les squamates. Les Rhynchocephalia, qui regroupe aujourd'hui les seuls Tuataras, étaient un groupe répandu et relativement prospère de lépidosauriens au début du mésozoïque, mais qui ont commencé à diminuer à partir du mi-Crétacé. Ils sont représentés aujourd'hui par un genre unique localisé exclusivement en Nouvelle Zélande.
L'ordre des squamates, qui est représenté aujourd'hui par les lézards, les serpents et les amphisbènes, a rayonné dans diverses niches écologiques pendant le jurassique et a réussi à passer au travers du crétacé. Ils ont survécu au passage de la limite K-T et sont actuellement le groupe le plus prospère et le plus diversifié de reptiles vivants avec plus de 6 000 espèces existantes. Aucune famille connue des squamates terrestres ne s'est éteinte à la limite, et les fossiles indiquent qu'ils n'ont souffert d'aucun déclin significatif dans leurs effectifs. Leur petite taille, leur métabolisme adaptable, et leur capacité à changer d'habitat pour trouver des conditions plus favorables ont été des facteurs clefs dans leur survie pendant la fin du crétacé et le début du paléocène.
Les reptiles marins non-archosaurien comprenant les mosasaures et les plésiosaures, les reptiles aquatiques géants qui étaient les prédateurs marins supérieurs, se sont éteints vers la fin du crétacé.
Archosaures
Le clade des archosauriens inclut deux ordres vivants, les crocodiliens (dont les Alligatoridae, les Crocodylidae et les Gavialidae sont les seules familles survivantes) et des dinosaures (dont les oiseaux sont les uniques membres survivants, alors que les dinosaures non-aviaires et les ptérosaures sont éteints).
Crocodiliens
Dix familles des crocodiliens ou de leurs proches parents sont déjà représentées dans les fossiles du Maastrichtien. Alors que cinq se sont éteintes avant la limite K-T, cinq autres familles ont des représentants fossiles jusqu'au Paléocène.
Toutes les familles de crocodiliens survivantes habitaient des environnements d'eau douce et terrestres, excepté les Dyrosauridae à la fois marins et dulçaquicole. La seule tendance apparente étant qu'aucun des grands crocodiles, tel que le crocodile nord-américain géant Deinosuchus, ne survécurent. La survie des crocodiliens à ces événements a pu simplement résulter de la persistance de leur niche aquatique et de leur capacité à creuser, qui a réduit leur susceptibilité aux effets négatifs sur l'environnement de la limite K-T.
En 2008, Stéphane Jouve et ses collègues ont suggéré que les juvéniles de Dyrosauridae auraient vécu dans l'eau douce comme les juvéniles des crocodiles marins modernes, ce qui les aurait aidés à survivre là où d'autres reptiles marins se sont éteints ; les environnements d'eau douce n'ayant pas été pas aussi fortement affectés par des événements de K-T que les environnements marins.
Ptérosauriens
La seule famille de ptérosauriens dont la présence au Maastrichtien est certaine, les Azhdarchidae, s'est éteinte à la limite K-T. Ces grands ptérosauriens étaient les derniers représentants d'un groupe en déclin qui comprenait dix familles durant le Crétacé moyen. Les ptérosauriens de plus petite taille s'étaient éteints avant le Maastrichtien au cours d'une période qui avait vu le déclin des espèces animales de petite taille au profit des espèces de grande taille. Simultanément, les oiseaux modernes avaient connu une forte diversification et avaient remplacé des oiseaux archaïques et des groupes de ptérosauriens, probablement en raison de la concurrence directe, ou simplement en remplissant des niches vides.
Dinosaures aviaires (Oiseaux)
La plupart des paléontologues considèrent les oiseaux comme les seuls survivants des dinosaures. Cependant, tous les oiseaux non-Néornithes se sont éteints, y compris des groupes florissants comme les enantiornithines et les hesperornithiforms. Plusieurs analyses de fossiles d'oiseaux montrent une divergence d'espèces avant la limite K-T, et que des parents du canard, du poulet et des autruches ont coexisté avec les dinosaures non-aviaires. Les oiseaux Néornithes ont survécu à la limite K-T peut-être en raison de leurs capacités de plonger, nager, ou chercher un abri dans l'eau et les marécages. Beaucoup d'espèces d'oiseaux peuvent construire les terriers, ou des nids dans les trous d'arbre ou les termite nests ⇔ termitières, ce qui les a mis à l'abri des effets sur l'environnement à la limite K-T. La survie à long terme après la limite a été garantie par la possibilité de remplir les nombreuses niches écologiques laissées vides par l'extinction des dinosaures non-aviaires.
Dinosaures non aviaires
L'extinction des dinosaures au passage de la limite K-T a donné lieu à plus de publications que n'importe quel autre groupe d'organismes. À l'exception de quelques revendications controversées, on convient que tous les dinosaures non-aviaires se sont éteints à la limite K-T. Les fossiles de dinosaures ont été interprétés à la fois pour montrer un déclin dans la diversité et aucun déclin dans la diversité pendant les derniers millions d'années du crétacé, et il se peut que la qualité des fossiles de dinosaures ne soit simplement pas assez bonne pour permettre à des chercheurs de distinguer les choix. Puisqu'il n'y a aucune preuve que les dinosaures non-aviaires de la fin du Maastrichtien aient pu creuser, nager ou plonger, ils ne pouvaient pas s'abriter pendant les plus mauvaises moments du stress environnemental qui se sont produites à la limite K-T. Il est possible que les petits dinosaures (autre que des oiseaux) aient survécu, mais ils auraient été privés de nourriture car des dinosaures herbivores auraient trouvé la matière végétale rare, et les carnivores se seraient rapidement trouvés à court de proies. Le consensus croissant au sujet du caractère endotherme des dinosaures (voir Physiology of dinosaurs (en)) aide à comprendre leur extinction complète par rapport à la survie de leurs parents proches, les crocodiliens. Les crocodiles étant ectothermes (animaux à « sang froid »), ils ont des besoins très limités en nourriture (ils peuvent survivre à plusieurs mois sans manger) tandis que des animaux de la taille semblable mais endothermes (à « sang chaud ») ont besoin de beaucoup plus de nourriture afin de soutenir leur métabolisme plus rapide. Ainsi, dans les circonstances de la rupture de chaîne alimentaire précédemment mentionnées, les dinosaures non-aviaires sont morts tandis que certains crocodiles survivaient. Dans ce contexte, la survie d'autres animaux endothermiques, tels que quelques oiseaux et mammifères, a pu être due, entre d'autres raisons, à leurs plus petits besoins de nourriture, liés à leur petite taille à l'époque de l'extinction.

Plusieurs chercheurs ont mentionné que l'extinction des dinosaures avait été progressive, de sorte qu'il y aurait eu des dinosaures au Paléocène (en). Ces arguments sont fondés sur la découverte de restes de dinosaures dans la Formation de Hell Creek (en) jusqu'à 1,3 mètres (4,27 ft) au dessus et 40 000 années après la limite K-T. Des échantillons de pollen prélevés près d'un fémur fossilisé d'hadrosaure découvert dans du grès à Ojo Alamo près du fleuve de San Juan indiquent que l'animal a vécu pendant le tertiaire, approximativement 64,5 MA (environ 1 million d'années après l'événement de K-T). Si leur existence après la limite K-T peut être confirmée, ces hadrosaures seraient considérés comme une clade mort-vivante. La recherche actuelle indique que ces fossiles ont été érodés de leurs endroits originaux et puis ré-ensevelis par des sédiments très postérieurs (retouchés).
Mammifères
Les groupes de mammifères existants aujourd'hui étaient déjà présents au Crétacé, qu'il s'agisse des monotrèmes qui pondent des œufs, des marsupiaux ou des placentaires, mais aussi d'autres groupes disparus comme les multituberculés, Dryolestoidea (en), et les Gondwanatheria (en). Tous ont survécu à l'événement de K-T, bien qu'ils aient enregistré des pertes. Beaucoup de marsupiaux ont disparu, en particulier ceux d'Amérique du Nord et plus particulièrement encore les espèces asiatiques regroupées dans le taxon des deltatheroïdes (en). Dans les gisements de fossiles de la formation de Hell Creek, au dessus de la limite K/T, on ne trouve plus de trace d'au moins la moitié des dix espèces de multituberculés ni d'aucune espèce marsupiale parmi les onze présentes avant la limite.
Les espèces de mammifères ont commencé à se diversifier approximativement 30 millions d'années avant la limite du Crétacé et du Tertiaire. Une radiation évolutive de mammifères s'est produite dans les quelques millions d'années qui ont suivi. La recherche actuelle indique que les mammifères n'ont pas eu d'explosion de diversification au passage de la limite K-T, en dépit des niches écologiques libérées par l'extinction des dinosaures. Plusieurs ordres de mammifères ont été interprétés comme se diversifiant juste après la limite K-T, comme les Chiroptera (chauves-souris) et les Cetartiodactyla (un groupe divers qui inclut aujourd'hui les baleines et dauphins et les Artiodactyla), mais des recherches plus récentes concluent que seuls les ordres de marsupaiux se sont diversifiés directement après la limite K-T.
Les espèces mammifères qui existaient à la limite K-T étaient généralement petites, de taille comparable aux rats ; cette petite taille les aurait aidées à trouver des abris dans des environnements protégés. En outre, on postule que quelques monotrèmes, marsupiaux, et placentaires primitifs étaient semi-aquatiques ou fouisseurs, car il existe encore de nombreuses lignées de mammifères ayant conservé de tels comportements aujourd'hui. Enfin, n'importe quel mammifère semi-aquatique ou creusant des terriers aurait eu la protection additionnelle contre le stress environnemental de la limite K-T.

















































