Cytoplasme - Définition
La liste des auteurs de cet article est disponible ici.
Peroxysome
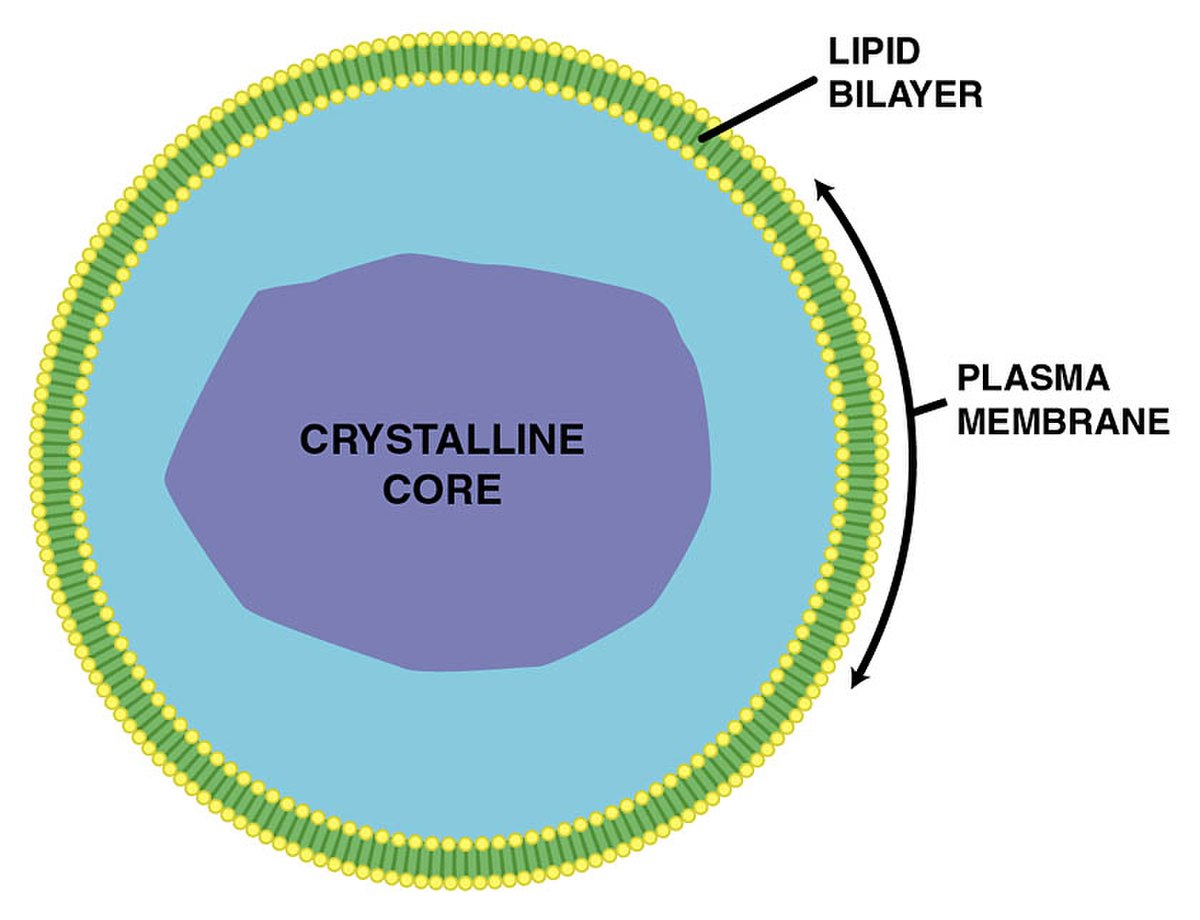
Les peroxysomes sont des microcorps à membrane sphérique, dont le diamètre est compris entre 0,5 et 1,5 micron. Ils se forment par gemmation à partir du réticulum endoplasmique lisse. Ces granules n'ont pas de structure interne. Ils possèdent un grand nombre d'enzymes métaboliques, en particulier l'enzyme catalase, qui catabolise la dégradation du peroxyde d'hydrogène (d'où provient le nom de peroxysome). La dégradation du peroxyde d'hydrogène est représentée par l'équation :

Ils effectuent des réactions d'oxydation qui ne produisent pas d'énergie directement utilisable par le reste de la cellule (ils ne génèrent pas d'ATP). À l'intérieur des peroxysomes est également dégradée la purine et, dans les plantes, elles sont impliquées dans la photorespiration.
Mitochondrie
La mitochondrie est un organite qui se trouve dans toutes les cellules eucaryotes, bien qu'elles puissent être absentes des cellules très spécialisées. Leur taille est généralement de cinq microns de large sur 0,2 µm d'épaisseur et possède une double membrane. L'espace inter-membranaire a une composition similaire à celle du hyaloplasme. Le rôle de cet espace est principalement de produire des molécules qui servent de précurseurs à la biosynthèse de macromolécules dans le hyaloplasme et d'être le lieu où se déroulent l'oxydation respiratoire et la synthèse des protéines mitochondriales. Cette dernière fonction se déroule de la même manière que la synthèse des protéines dans le hyaloplasme.
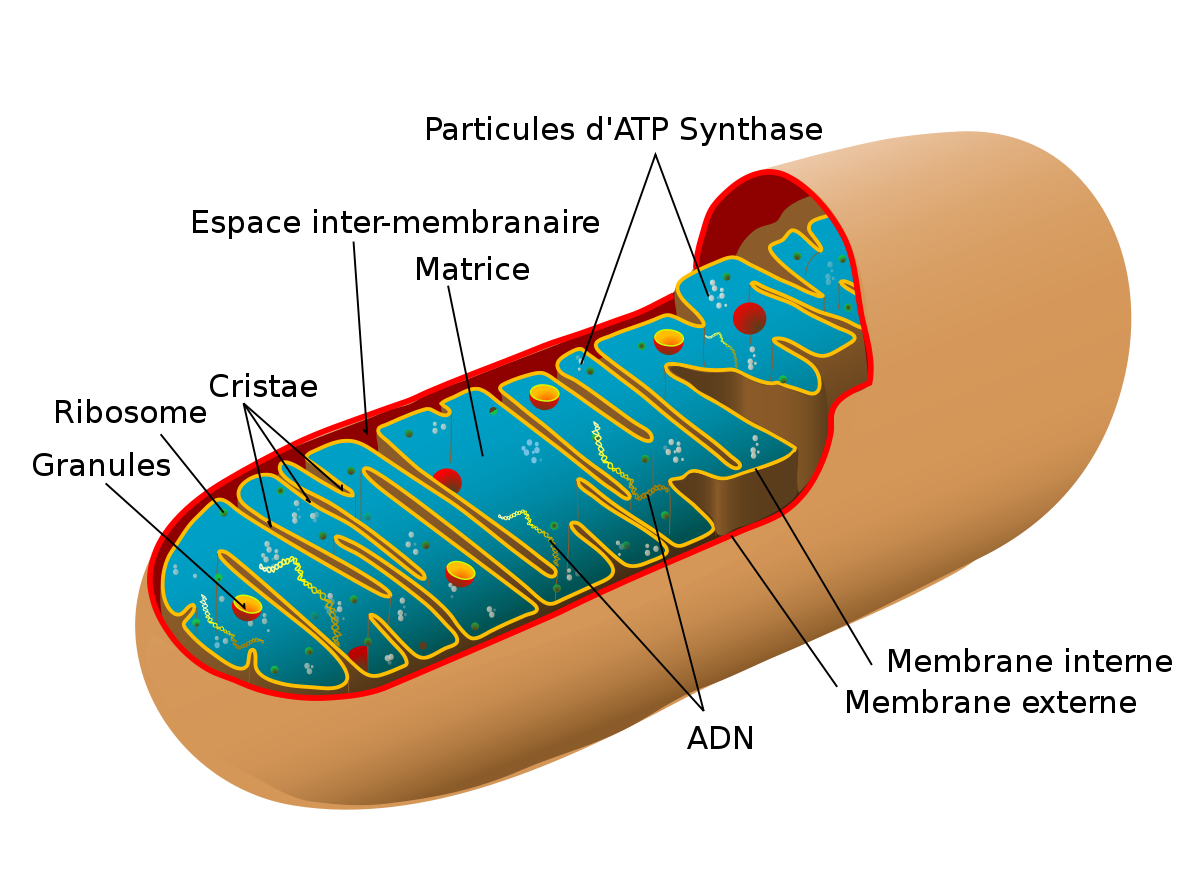
Les membranes de la mitochondrie sont constituées de phospholipides et de protéines, qui se combinent pour former un réticulum lipido-protéiné. La membrane la plus externe contrôle l'entrée et la sortie de substances dans la cellule et sépare l'organite du hyaloplasme. La membrane externe contient des protéines de transport spécialisées qui permettent le passage de molécules du cytosol jusqu'à l'espace inter-membranaire. Ces protéines forment de grands canaux d'eau ou de porines, qui rendent la membrane très perméable, contrairement à la membrane interne. La membrane est également constituée d'enzymes comme ceux qui activent l'oxydation des acides gras dans la matrice.
La membrane interne est repliée vers le centre de la mitochondrie ce qui donne lieu à des extensions, les crêtes mitochondriales ou encore cristae. Ceux-ci s'étendent sur toute la largeur de l'organite et présentent des granules à leur surface. De facto, c'est donc la membrane qui possède la plus grande surface, en raison de ces crêtes mitochondriales. C'est également la membrane cellulaire la plus riche en protéines. On y trouve notamment : les protéines qui forment la chaîne de transport des électrons vers l'oxygène moléculaire (chaîne respiratoire), un complexe enzymatique appelé ATP synthase, qui agit comme catalyseur lors de la synthèse de l'ATP, ainsi que des protéines de transport qui permettent le passage des ions et des molécules à travers la membrane mitochondriale interne. Cette membrane présente l'avantage de ne pas avoir de cholestérol entre ses lipides et d'être riche en un phospholipide rare, la cardiolipine. D'une façon générale, sa fonction principale est d'être le lieu du processus respiratoire.
La mitochondrie se divise donc en deux espaces distincts : l'espace inter-membranaire et la matrice. La matrice a une consistance de gel en raison de la présence d'une concentration élevée de protéines hydrosolubles et de 50% d'eau. Par ailleurs, elle contient des molécules d'ADN (l'ADN mitochondrial) qui contiennent l'information pour synthétiser un bon nombre de protéines mitochondriales, des molécules d'ARN mitochondrial formant les ribosomes mitochondriaux (ou mitoribosomes), des ions, du calcium, des phosphates, de l'ADP, de l'ATP, des coenzymes-A et une grande quantité d'enzymes.
Au final, la mitochondrie permet non seulement l'oxydation du pyruvate en CO2 couplée à la réduction des porteurs électroniques NAD+ et FAD (NADH et FADH2), mais également le transfert des électrons depuis le NADH et le FADH2 vers le dioxygène couplé à la génération d'une force protomotrice, ou encore l'utilisation de l'énergie stockée dans le gradient électrochimique de protons pour la synthèse de l'ATP du complexe FO-F1.

















































