Contraction musculaire - Définition
La liste des auteurs de cet article est disponible ici.
Motoneurone
Les motoneurones sont les neurones qui contrôlent les fibres musculaires.
Myocyte ou fibre musculaire
Un myocyte ou fibre musculaire est une cellule musculaire de forme très allongée dont les extrémités sont constituées de filaments de collagène. Chaque fibre musculaire est en contact avec une fibre nerveuse qui commande son activité. La fibre musculaire a deux propriétés fondamentales, l'excitabilité sous l'action stimulatrice de la fibre nerveuse, et la contractilité, résultat ultime de la stimulation. Lorsqu'une fibre musculaire se contracte, sa longueur diminue, ce qui génère un mouvement de rapprochement de ses extrémités. Si l'on raisonne au niveau du corps musculaire en prenant le biceps comme exemple, la stimulation de toutes les fibres musculaires provoque la contraction du muscle ce qui se traduit par un rapprochement de ses deux extrémités. Comme l'une de ses extrémités est attachée à l'avant-bras, le raccourcissement du biceps provoque, de part et d'autre de l'articulation du coude, la flexion de l'avant-bras.
Myofibrille
Les myofibrilles sont les fibres contractiles, actine et myosine, localisées à l'intérieur de la cellule musculaire.
Sarcolemme
Un sarcolemme est une structure histologique composée de la membrane plasmique des cellules musculaires et de la lame basale qui entoure ces mêmes cellules.
La contraction
Il existe deux modalités de contraction musculaire :
- La contraction isométrique lorsqu'il n'y a pas de déplacement osseux ;
- La contraction anisométrique quand il y a déplacement des pièces osseuses, avec un rapprochement des points d'insertion des muscles elle est concentrique et excentrique quand ceux-ci s'éloignent.
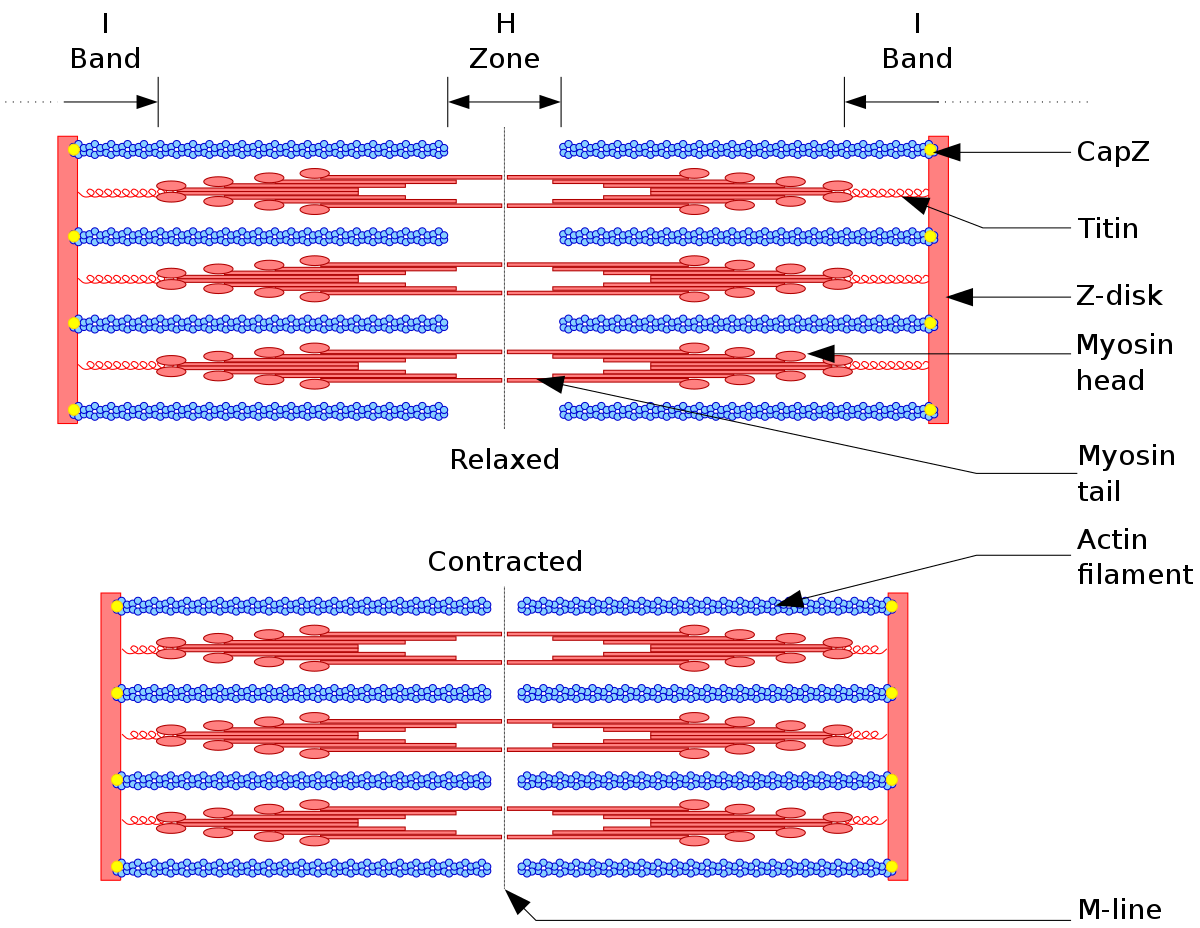
Les protéines contractiles
Les protéines contractiles sont des protéines dont la déformation spatiale est responsable d’un mouvement de la cellule. Dans la fibre musculaire striée squelettique, les protéines contractiles sont la myosine et l'actine.
Fixation du calcium
La fixation de 4 molécules de calcium au niveau de la troponine C, permet l'activation (plutôt une levée d'inhibition) de la mise en place des ponts actine-myosine d'où la contraction.
Le couplage excitation-contraction
Le couplage excitation-contraction est l’ensemble des phénomènes qui permettent le passage du signal depuis le sarcolemme vers les myofibrilles. Au repos, la concentration du calcium dans le cytoplasme des fibres musculaires est très basse. L’arrivée du potentiel d’action musculaire au niveau de structures spécialisées que sont les triades va provoquer un flux de calcium depuis le réticulum sarcoplasmique vers le cytoplasme de la cellule.
Le tubule T

Le tubule T (pour tubule transverse), est une invagination du sarcolemme, permettant de conduire le potentiel d’action musculaire jusqu’au cœur de la cellule. Les tubules T sont répartis régulièrement au sein de la cellule et sont positionnés à la jonction bande A/bande I.
La membrane du tubule T contient une des trois protéines clé du couplage excitation contraction : le récepteur de la dihydropyridine (DHP). Cette protéine est sensible à la variation de voltage résultant du potentiel d’action musculaire.
Signalons tout de suite que le nom de récepteur de la dihydropyridine (comme celui de récepteur de la ryanodine au paragraphe suivant), n’a rien à voir avec la fonction physiologique de cette protéine mais est lié à sa découverte et à ses propriétés pharmacologiques (cf paragraphe sur la pharmacologie).
Le réticulum sarcoplasmique
Le réticulum sarcoplasmique est un compartiment intracellulaire spécialisé dans le stockage et la libération du calcium. C’est ce haut niveau de spécialisation du réticulum dans les fibres musculaires qui a amené à la dénomination spécifique de « réticulum sarcoplasmique », alors que ce compartiment est appelé « réticulum endoplasmique » pour les autres types cellulaires.
La structure est très particulière et on peut distinguer deux parties : les citernes terminales, gros renflements situées de part et d’autre des tubules T, et le réticulum sarcoplasmique longitudinal, fin réseau reliant les citernes terminales. Cette différence structurale est assortie d’une différence fonctionnelle, les citernes terminales étant plus spécialement impliquées dans la libération du calcium et le réticulum sarcoplasmique longitudinal dans sa recapture ou repompage.
La membrane des citernes terminales contient la deuxième protéine clé du couplage excitation contraction : le récepteur à la ryanodine. Cette protéine est un canal calcique qui permet, lorsqu’il s’ouvre, le passage du calcium du réticulum sarcoplasmique vers le cytoplasme.
La membrane du réticulum sarcoplasmique longitudinal contient la troisième protéine clé : la calcium ATP-ase ou SERCA. Cette enzyme est une pompe permettant l’accumulation du calcium à l’intérieur du réticulum, grâce à l’énergie fournie par l’hydrolyse de l’ATP.
La triade
L’association d’un tubule T et des deux citernes terminales adjacentes forme une structure caractéristique appelée triade. La membrane du tubule T est à proximité du réticulum sarcoplasmique. Chaque récepteur à la dihydropyridine (dans la membrane du tubule T) est associé avec un récepteur à la ryanodine (dans la membrane du réticulum).
La libération du calcium
La variation du potentiel de membrane induite par l’arrivée du potentiel d’action musculaire induit un changement conformationnel du récepteur à la dihydropyridine ; c’est l’élément voltage-sensible. Ce changement conformationnel induit à son tour l’ouverture du récepteur à la ryanodine associé, ce qui permet au calcium de sortir du réticulum sarcoplasmique dans le cytoplasme. Ce qui permet par l'intermédiaire des kinétochore d'activer les microtubules membranaires ancrés à la bicouche lipidique grâce a un domaine hydrophobe en hélice alpha.

















































