Cladistique - Définition
La liste des auteurs de cet article est disponible ici.
Concepts importants de la cladistique
Le clade comme unité de classification
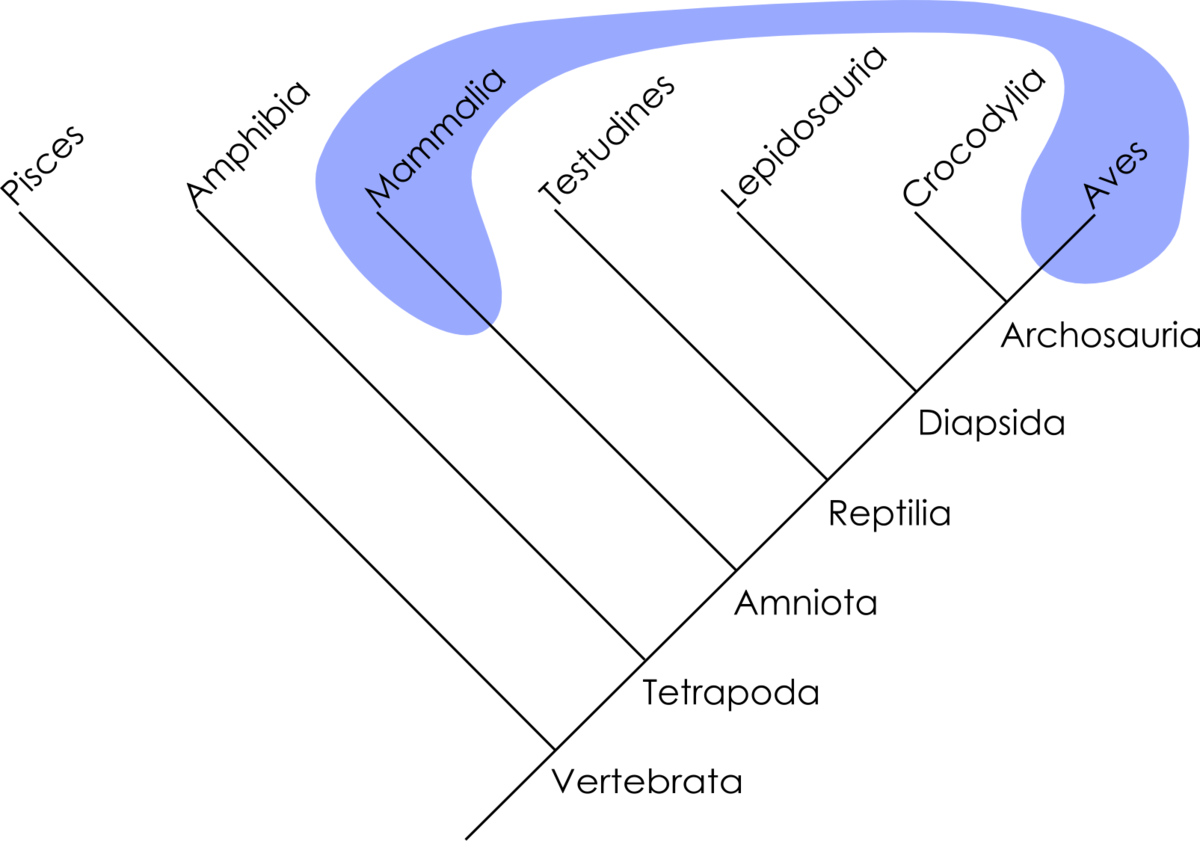
La cladistique reconnait une unité de classement bien particulière, le clade. Un clade est un groupe de taxons formant un groupe monophylétique, une totalité de descendance, un ancêtre commun et tous ses descendants. Ce concept s'oppose à celui du grade, rapprochement de taxons reposant sur d'autres critères (e.g. ressemblance générale, somme de modifications adaptatives). La cladistique qualifiera souvent ces grades de paraphylétiques ou polyphylétiques selon que le rapprochement soit effectué sur la base de plésiomorphies ou d'homoplasies. Certains grades sont cependant monophylétiques. Par exemple, les algues forment un grade polyphylétique, les reptiles forment un grade paraphylétique et les mammifères forment un grade monophylétique.
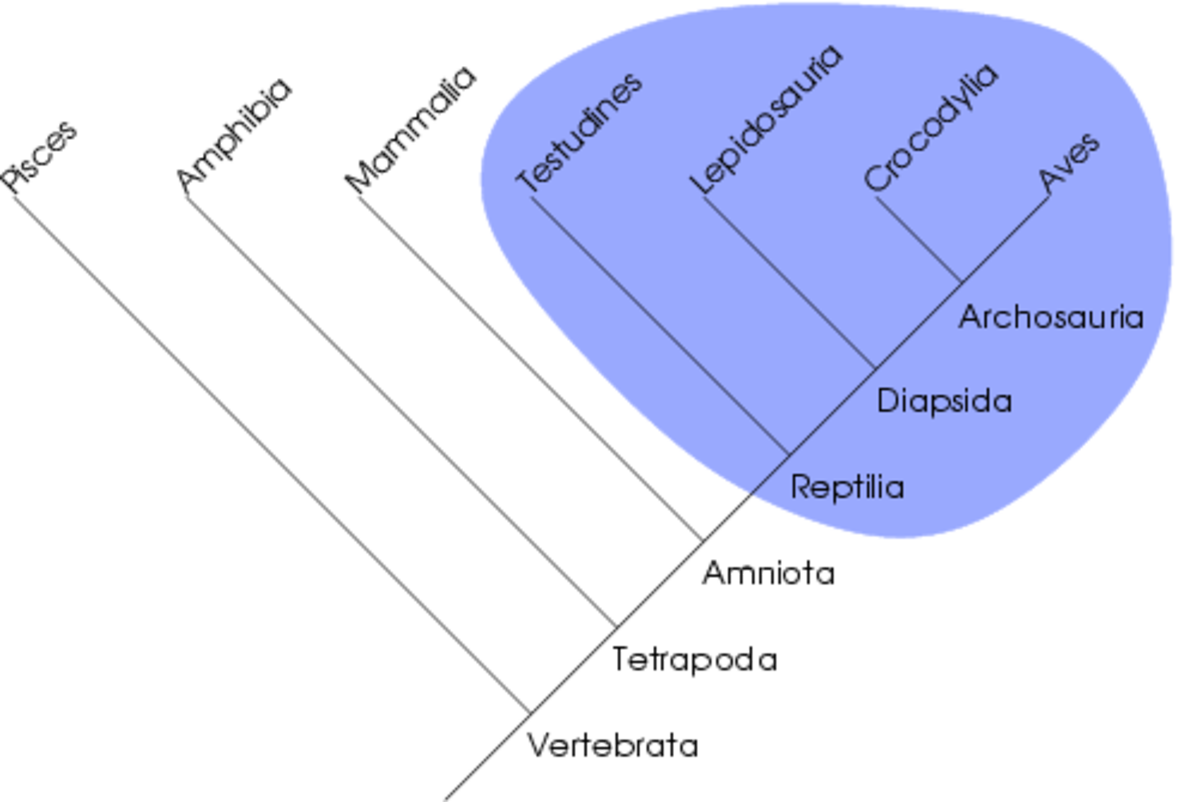
La notion de clade peut aussi être définie par un ensemble de taxons plus proches qu'ils ne le sont entre eux que de tout autre taxon. Par exemple, les reptiles ne forment pas un clade car certains sont plus apparentés aux oiseaux qu'aux autres reptiles (les Crocodiliens sont inclus dans les Archosauriens comme les oiseaux). Il est à noter qu'un clade représente uniquement une unité évolutive. On parle aussi parfois de groupe holophylétique, ce qui est synonyme de groupe monophylétique.
Paraphylie
Si on parle de groupe monophylétique pour une totalité de descendance, la notion de paraphylie s'applique à un groupe défini sur un ou des caractères plésiomorphes. Ce n'est donc pas une totalité de descendance puisque les organismes portant les états dérivés en sont exclus. Un groupe paraphylétique n'est donc pas monophylétique. Il est donc important en cladistique de bien séparer caractère plésiomorphe et apomorphe pour déterminer si un groupe est monophylétique ou non. La méthode cladistique ne permet donc pas l'existence de groupes tel que les reptiles, les poissons, les invertébrés, les procaryotes, etc. Dans un groupe paraphylétique, certains taxons sont plus proches d'autres taxons hors du groupe que de taxons au sein du groupe paraphylétique. Dans l'exemple des poissons, si on considère les Tétrapodes comme groupe dérivé et les poissons comme groupe primitif par rapport aux Tétrapodes, on constate que certains poissons sont plus proches des Tétrapodes qu'ils ne le sont d'autres poissons : par exemple le Cœlacanthe est plus proche des Tétrapodes que du requin.
La notion de polarisation et de sens de transformation est ici importante. Un caractère pouvant exister sous deux états: primitif (plésiomorphe) ou dérivé (apomorphe), il faut pouvoir déterminer quel état est apomorphe et lequel est plésiomorphe. Pour cela on peut utiliser plusieurs critères dont le plus utilisé est celui du groupe externe. D'autres critères sont utilisés comme le critère ontogénétique et à moindre mesure les critères paléontologique et chorologique. Soient deux états a et b d'un même caractère ; on ne connait pas le sens de transformation entre a et b on a donc a↔b. On peut donc admettre deux sens de transformation : a→b ou b→a. Dans le cas a→b, a se transforme en b, donc a est plésiomorphe par rapport à b et seul b nous renseigne sur les relations de parenté. Dans le cas b→a, b se transforme en a, b est donc plésiomorphe par rapport à a et seul a nous renseigne sur les relations de parenté.
En cladistique un ensemble d'espèces ne peut pas être primitif : il n'y a pas de groupe ancestral. Si un clade contient plusieurs groupes, c'est que chacun de ces groupes a des caractères dérivés qui les distinguent des autres (autapomorphies), ce qui implique qu'un groupe ancestral ne peut pas exister en cladistique. La notion d'ancêtre pourrait s'appliquer à l'espèce, cependant une espèce ancestrale est nécessairement définie par des caractères primitifs donc plésiomorphes par rapport à ses descendants, elle est donc forcément paraphylétique. En cladistique l'ancêtre est donc une hypothèse et n'est pas identifié, ce qui, évidemment, n'implique pas qu'il n'existe pas mais simplement qu'il ne peut pas être défini en tant que taxon ou espèce mais simplement en tant qu'hypothèse d'un ensemble de caractères primitifs (on parle alors de morphotype ancestral).
Homologie vs. homoplasie
On distingue deux types de ressemblances, l'homologie et l'homoplasie. Des caractères homologues sont hérités d'un ancêtre commun, la relation liant deux homologues étant appelée homologie. À l'inverse, l'homoplasie, terme introduit par Lankester en 1870, caractérise des ressemblances entre caractères n'étant pas hérités d'un ancêtre commun. Différencier l'homoplasie de l'homologie est une tâche compliquée et plusieurs critères définissent l'homologie dont le plus utilisé est le principe des connexions d'Étienne Geoffroy Saint-Hilaire appelé aussi identité des connexions. Ce principe sera repris par Richard Owen. d'après ce principe, sont homologues deux organes si quelles que soient leurs formes ou fonctions, ils ont les même connexions avec d'autres organes. Ce principe peut aisément s'étendre aux séquences moléculaires et l'alignement de séquences correspond à l'application de ce principe.
En réalité ce principe ne permet que de formuler des hypothèses dites d'homologie primaire. En effet rien ne nous assure que le caractère supposé homologue l'est effectivement et que ce n'est pas de l'homoplasie tant que l'hypothèse d'homologie n'est pas testée. Le test effectué est le test de congruence selon Hennig ou de parcimonie. En cladistique on cherche à minimiser le nombre d'hypothèses surnuméraires ou ad hoc. Une hypothèse ad hoc en reconstruction phylogénétique étant l'apparition indépendante d'un caractère dans deux lignées différentes, c'est-à-dire de l'homoplasie. À l'inverse, l'hypothèse de base est que le même caractère observé chez deux taxons indique une relation de parenté. En maximisant alors l'homologie (ou en minimisant l'homoplasie, ce qui constitue deux démarches équivalentes), on obtiendra un test des hypothèses primaires d'homologie. On retiendra alors le cladogramme maximisant l'homologie. Les caractères apparaissant une fois dans le cladogramme résultant de l'analyse de parcimonie seront considérés comme effectivement homologues et donc hérités d'un ancêtre commun. L'homologie secondaire n'est donc plus seulement une homologie déduite du principe des connexions mais une homologie de parenté ou de descendance.
À l'inverse, certains caractères peuvent se révéler homoplastiques une fois l'arbre reconstitué. Ces caractères impliquent au moins deux pas évolutifs ou deux hypothèses de transformation dans le cladogramme résultant. L'homoplasie est donc une hypothèse d'homologie primaire rejetée. Il existe en fait deux types d'homoplasie: la Convergence évolutive et la réversion, décelables entre elles seulement sur l'arbre enraciné le plus parcimonieux. La convergence indique que le même caractère (au sens des connexions et non au sens évolutif) est apparu au moins deux fois dans le groupe considéré. À l'inverse, la réversion est la perte secondaire d'un caractère, c'est-à-dire le retour à l'état primitif. Par exemple l'état primitif des membres pairs chez les Vertébrés est l'absence. L'état dérivé chez les Gnathostomes est leur présence. Or ce caractère est perdu chez les Gymnophiones ou les Serpents. Une réversion est une homoplasie qui s'effectue sur une même morphocline (suite de transformations) alors que la convergence s'effectue sur plusieurs morphoclines distinctes. L'homoplasie n'étant pas un caractère hérité par un ancêtre commun, elle ne nous renseigne pas sur les relations de parenté. Un groupe formé sur une homoplasie est appelé groupe polyphylétique.
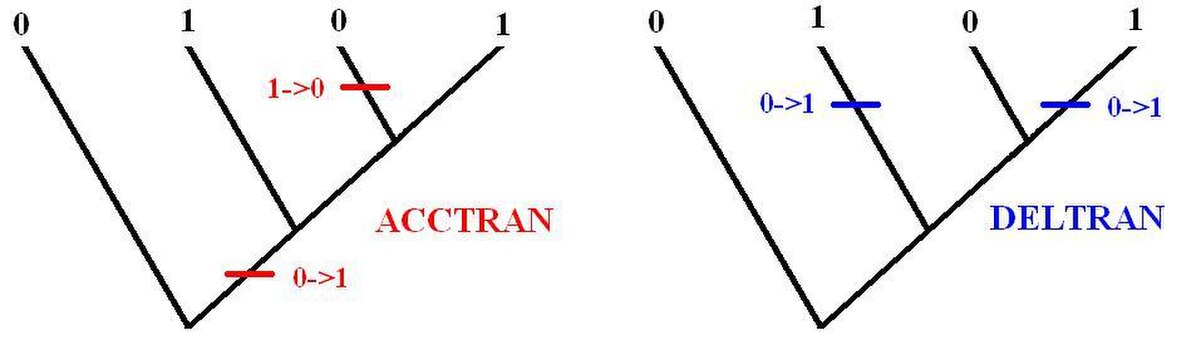
Dans certains cas il n'y a aucun élément pour savoir si l'homoplasie est une réversion ou une convergence, on parle alors de synapomorphie ambigüe. À partir du cladogramme, le choix de savoir si le caractère est issu d'une convergence ou d'une réversion est arbitraire. L'hypothèse ACCTRAN (Accelerated transformation) suppose des réversions. L'hypothèse DELTRAN (Delayed transformation) suppose des convergences. Si le choix de l'un ou l'autre n'est pas important en moléculaire, le choix est bien moins anodin pour les phylogénies morphologiques car le choix de l'une ou l'autre des hypothèses change l'interprétation des caractères. Cependant le choix de l'un ou l'autre n'influence pas les relations de parentés. L'image ci dessous l'illustre pour deux arbres ayant la même répartition des caractères, on a deux configurations des transformations différentes.