Chaîne respiratoire - Définition
La liste des auteurs de cet article est disponible ici.
Théorie chimio-osmotique de Mitchell
Il existe 5 ensembles de protéines et de coenzymes impliqués dans les oxydations phosphorylantes de la chaîne respiratoire. Les 4 premiers complexes (I, II, III et IV) interviennent dans le transport des électrons et le cinquième (V) intervient dans la synthèse d'ATP (figure 2). Ces complexes diffusent de façon indépendante au sein de la membrane interne et connectés par un transporteur liposoluble mobile le coenzyme Q (CoQ) et le cytochrome C fixé à la membrane.
- Complexe I : NADH-ubiquinone réductase ;
- Complexe II : Succinate-ubiquinone réductase ;
- Complexe III : Ubiquinone-cytochrome C réductase ;
- Complexe IV : Cytochrome oxydase ;
- Complexe V : ATP synthase.
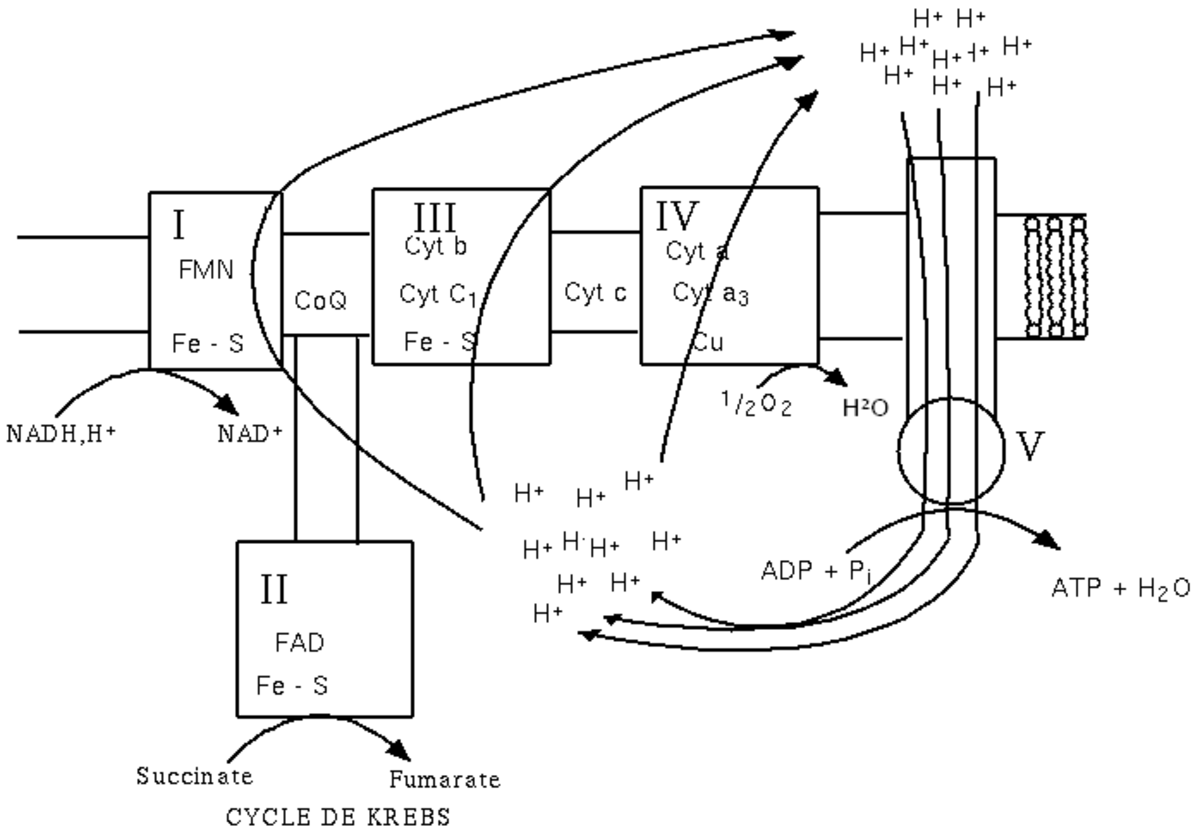
L'expulsion d'ions H+ entraîne un gradient de pH. Les ions H+ vont revenir (dans la sens du gradient électrochimique) dans la matrice mitochondriale (ou dans le cytoplasme des bactéries) en entraînant un flux d'ions dans :
- L'ATP synthase (complexe V) entraînant la synthèse d'ATP ;
- Pour le transport de molécules contre un gradient de concentration (exemple : pompe à sodium) ;
- Pour la rotation des flagelles bacteriens.
Régulation
La régulation de la chaîne respiratoire dépend des apports en:
- NADH,H+
- O2
- ADP+Pi
Au repos, le rapport :
![\frac {[ATP]}{[ADP+Pi]}](https://static.techno-science.net/illustrationWebp/Definitions/autres/0/0b47cc09d09bc275e82324750a69a513_d0fc224c55c5c777dbd6c12e92143d5a.png)
Pendant un effort :
Transport des molécules d'ATP formées
Les molécules d'ATP ainsi formées se retrouvent dans la matrice mitochondriale où leur présence est peu intéressante. Pour passer dans le hyaloplasme, elles empruntent un anti-port, une protéine permettant de faire passer de l'ATP dans le sens Matrice mitochondriale (ANT1 ou 3)→ espace intermembranaire → cytosol et de l'ADP dans le sens inverse.
La différence de potentiel crée en partie par le gradient de protons induit un transport 30 fois plus rapide de l'ATP par rapport à l'ADP.
Complexes et fonctionnement
- Complexe II : Livraison d'électrons de plus faible énergie provenant du succinate à l'ubiquinone :
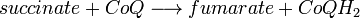

- Complexe III :
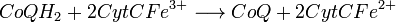

De plus, on assiste au transfert de quatre protons (H + ). On peut remarquer aussi que le cytochrome c (CytC) est très conservé dans l'évolution des espèces.
- Complexe IV :
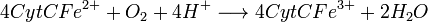

- Complexe V : Dernière étape:
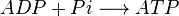

Les trois sous-unités β ont, à un moment donné une forme différente, en accord avec une activité fonctionnelle différente. Elles peuvent être :
- O (Open)
- L (Loose) → Faible affinité pour le substrat
- T (Tight) → Forte affinité pour le substrat
Des chaînes respiratoires particulières : celles des bactéries
- La chaîne respiratoire des bactéries dites « oxydase + » (Pseudomonas, Vibrio...) est sensiblement la même que celle des mitochondries.
- La chaîne respiratoire des bactéries dites « oxydase - » (Enterobacteriaceae...) est plus courte : il n'y a pas le complexe IV de la chaîne respiratoire correspondant à la cytochrome oxydase.
- Dans certains cas (respirations anaérobies chez les bactéries), l'accepteur final de la chaîne respiratoire n'est pas du dioxygène mais un composé organique comme du CO. Voir respiration anaérobie.

















































