Bacteria - Définition
La liste des auteurs de cet article est disponible ici.
Mobilité des bactéries
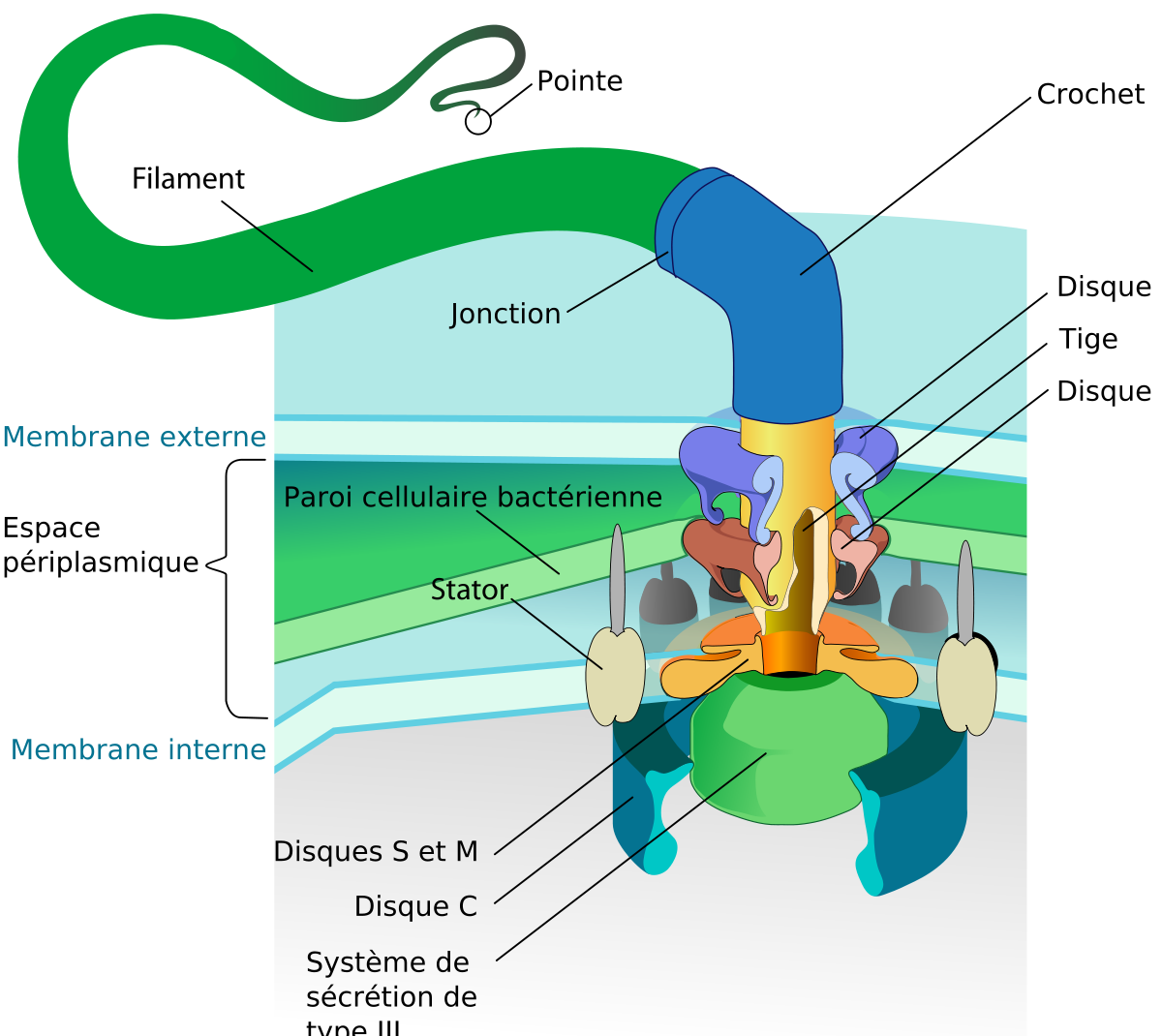
Certaines bactéries sont mobiles et peuvent se déplacer grâce à un ou plusieurs flagelles, d’autres bactéries peuvent se déplacer par glissement.
Les flagelles des bactéries sont de longs appendices protéiques flexibles. Leur nombre et leur position peuvent différer selon les espèces de bactéries. La flagellation (ou ciliature) polaire monotriche correspond à la présence d’un seul flagelle à un pôle de la bactérie (exemple des Vibrio). La flagellation polaire lophotriche correspond à la présence de plusieurs flagelles au pôle de la bactérie (Pseudomonas par exemple). D’autres bactéries comme Escherichia coli produisent des flagelles sur toute la surface cellulaire et possèdent donc une flagellation péritriche.
Beaucoup de bactéries (comme Escherichia coli) ont deux modes distincts de circulation : mouvement vers l'avant (natation) et mouvement de rotation ou mouvement de « roulis » également et appelé « lang » en anglais. Le tumbling leur permet de se réorienter et leur fait faire un mouvement en trois dimensions, évoluant vers un mouvement de marche au hasard. Les flagelles d'un groupe de bactéries, les spirochaetes, se trouvent entre deux membranes dans l'espace périplasmique.
Le filament du flagelle est constitué d’une protéine, la flagelline. Le type de rotation du flagelle peut déterminer le type de mouvement de la bactérie.
Les bactéries mobiles peuvent réagir à des stimuli, être attirées par des substances nutritives comme les sucres, les acides aminés, l’oxygène, ou être repoussées par des substances nuisibles. il s'agit de la chimiotaxie, phototaxis et magnetotaxis. Ce comportement est appelé le chimiotactisme. Des chimiorécepteurs de nature protéique sont présents au niveau de la membrane plasmique et du périplasme des bactéries et peuvent détecter différentes substances attractives ou nocives.
Les ions de cuivre bloquent la rotation des flagelles. Pour le faire repartir, on recourt à l’acide éthylènediaminetétraacétique, capable de capturer les ions et donc d'en libérer le flagelle.
Interactions avec d’autres organismes
En dépit de leur apparente simplicité, les bactéries peuvent entretenir des associations complexes avec d’autres organismes. Ces associations peuvent être répertoriées en parasitisme, mutualisme et commensalisme. En raison de leurs petites tailles, les bactéries commensales sont ubiquitaires et sont rencontrées à la surface et à l’intérieur des plantes et des animaux.
Mutualistes
Dans le sol, les bactéries de la rhizosphère (couche de sol fixée aux racines des plantes) fixent l’azote et produisent des composés azotés utilisés par les plantes (exemple de la bactérie Azotobacter ou Frankia). En échange, la plante excrète au niveau des racines des sucres, des acides aminés et des vitamines qui stimulent la croissance des bactéries. D’autres bactéries comme Rhizobium sont associées aux plantes légumineuses au niveau de nodosités sur les racines.
Il existe de nombreuses relations symbiotiques ou mutualistes de bactéries avec des invertébrés. Par exemple, les animaux qui se développent à proximité des cheminées hydrothermales des fonds océaniques comme les vers tubicoles Riftia pachyptila, les moules Bathymodiolus ou la crevette Rimicaris exoculata vivent en symbiose avec des bactéries chimiolitho-autotrophes.
Buchnera est une bactérie endosymbiote des aphides (puceron). Elle vit à l'intérieur des cellules de l'insecte et lui fournit des acides aminés essentiels. La bactérie Wolbachia est hébergée dans les testicules ou les ovaires de certains insectes. Cette bactérie peut contrôler les capacités de reproduction de son hôte.
Des bactéries sont associées aux termites et lui apportent des sources d'azote et de carbone.
Des bactéries colonisant la panse des herbivores permettent la digestion de la cellulose par ces animaux. La présence de bactéries dans l’intestin de l’Homme contribue à la digestion des aliments mais les bactéries fabriquent également des vitamines comme l’acide folique, la vitamine K et la biotine.
Des bactéries colonisent le jabot d'un oiseau folivore (consommateur de feuilles), le Hoazin (Opisthocomus hoazin). Ces bactéries permettent la digestion de la cellulose des feuilles, de la même manière que dans le rumen des ruminants.
Des bactéries bioluminescentes comme Photobacterium sont souvent associées à des poissons ou des invertébrés marins. Ces bactéries sont hébérgées dans des organes spécifiques chez leurs hôtes et émettent une luminescence grâce à une protéine particulière : la luciférase. Cette luminescence est utilisée par l'animal lors de divers comportements comme la reproduction, l'attraction de proies ou la dissuasion de prédateurs.
Pathogènes
pour l'Homme
Les bactéries pathogènes sont responsables de maladies humaines et causent des infections. Les organismes infectieux peuvent être distingués en trois types : les pathogènes obligatoires, accidentels ou opportunistes.
Un pathogène obligatoire ne peut survivre en dehors de son hôte. Parmi les bactéries pathogènes obligatoires, Corynebacterium diphtheriae entraîne la diphtérie, Treponema pallidum est l’agent de la syphilis, Mycobacterium tuberculosis provoque la tuberculose, Mycobacterium leprae la lèpre, Neisseria gonorrhoeae la gonorrhée. Les Rickettsia à l’origine du typhus sont des bactéries parasites intracellulaires.
Un pathogène accidentel présent dans la nature peut infecter l’Homme dans certaines conditions. Par exemple, Clostridium tetani provoque le tétanos en pénétrant dans une plaie. Vibrio cholerae entraîne le choléra suite à la consommation d’une eau contaminée.
Un pathogène opportuniste infecte des individus affaiblis ou atteints par une autre maladie. Des bactéries comme Pseudomonas aeruginosa, des espèces de la flore normale, comme des Staphylococcus de la flore cutanée, peuvent devenir des pathogènes opportunistes dans certaines conditions. On rencontre ce type d’infection surtout en milieu hospitalier.
La capacité d’une bactérie à provoquer une maladie est son pouvoir pathogène. L’intensité du pouvoir pathogène est la virulence. L’aboutissement de la relation bactérie-hôte et l’évolution de la maladie dépendent du nombre de bactéries pathogènes présentes dans l’hôte, de la virulence de cette bactérie, des défenses de l’hôte et de son degré de résistance.
Pour déclencher une maladie, les bactéries infectieuses doivent d’abord pénétrer dans l’organisme et adhérer à un tissu. Des facteurs d’adhésion permettent la fixation des bactéries à une cellule. Le pouvoir invasif est la capacité de la bactérie à se répandre et à se multiplier dans les tissus de l’hôte. Les bactéries peuvent produire des substances lytiques lui permettant de se disséminer dans les tissus. Certaines bactéries présentent aussi un pouvoir toxinogène qui est la capacité de produire des toxines, substances chimiques portant préjudice à l’hôte. On peut distinguer les exotoxines libérées lors de la multiplication des bactéries et les endotoxines fixées dans la membrane des bactéries.
Les bactéries pathogènes tentant d’envahir un hôte rencontrent toutefois de nombreux mécanismes de défense assurant à l’organisme une protection aux infections. Une bonne alimentation et une hygiène de vie correcte constituent une première protection. La peau, les muqueuses forment une première ligne de défense contre la pénétration d’organismes pathogènes. Les bactéries de la flore normale constituent aussi une barrière de protection. Lorsqu’un micro-organisme a pénétré ces premières lignes de défense, il rencontre des cellules spécialisées qui se mobilisent contre l’envahissement : ce sont les phagocytes. L’inflammation est une réaction défensive non spécifique. Un second système de défense très efficace est le système immunitaire spécifique, capable de reconnaître des antigènes portés ou sécrétés par les bactéries, et d’élaborer des anticorps et des cellules immunitaires spécifiques de ces antigènes.
pour les plantes
Les bactéries pathogènes pour les plantes sont connues du grand public pour leur responsabilité dans la dévastation de cultures agricoles. En 2001, les vergers du midi de la France étaient victimes d'une vague d'infection par une bactérie du genre xanthomonas
En biotechnologie végétale, la bactérie du sol Agrobacterium tumefaciens est utilisée pour sa capacité à transmettre un fragment d'ADN à la plante cible lors de son cycle infectieux.

















































