Acide ribonucléique - Définition
La liste des auteurs de cet article est disponible ici.
Introduction


L'acide ribonucléique, ou ARN, est une molécule biologique trouvée dans pratiquement tous les organismes vivants, y compris certains virus. L'ARN est une molécule très proche chimiquement de l'ADN et il est d'ailleurs en général synthétisé dans les cellules à partir d'une matrice d'ADN dont il est une copie. Les cellules vivantes utilisent en particulier l'ARN comme un support génétique intermédiaire de nos gènes pour fabriquer les protéines dont elles ont besoin. L'ARN peut remplir de nombreuses autres fonctions et en particulier intervenir dans des réactions chimiques de la cellule.
Chimiquement, l'ARN est un polymère linéaire constitué d'un enchaînement de nucléotides. Chaque nucléotide contient un groupement phosphate, un sucre, le ribose et une base azotée. Les nucléotides sont liés les uns aux autres par des liaisons phosphodiester. On trouve quatre bases azotées dans l'ARN, l'adénine, la guanine, la cytosine et l'uracile. L'ARN a de nombreuses similarités avec l'ADN, avec cependant quelques différences importantes : sur le plan de la structure, l'ARN contient un ribose à la place du désoxyribose de l'ADN, ce qui rend l'ARN chimiquement plus instable et la thymine de l'ADN y est remplacée par l'uracile, qui possède les mêmes propriétés d'appariement de base avec l'adénine. Sur le plan fonctionnel, l'ARN est le plus souvent trouvé dans les cellules sous forme de simple brin, tandis que l'ADN est présent sous forme de deux brins complémentaires, formant une double hélice. Enfin les molécules d'ARN trouvées dans les cellules sont plus courtes (de quelques dizaines à quelques milliers de nucléotides) que l'ADN du génome (de quelques millions à quelques milliards de nucléotides).
Dans la cellule, l'ARN est produit par transcription à partir de l'ADN situé dans le noyau. L'ARN est donc une copie d'une région de l'un des brins de l'ADN. Les enzymes qui effectuent cette copie ADN→ARN s'appellent des ARN polymérases. Les ARN ainsi produits peuvent avoir trois grands types de fonctions, ils peuvent être support de l'information génétique d'un ou plusieurs gènes codant pour des protéines (on parle alors d'ARN messagers), ils peuvent adopter une structure secondaire et tertiaire stable et accomplir des fonctions catalytiques (par exemple l'ARN ribosomique), ils peuvent enfin servir de guide ou de matrice pour des fonctions catalytiques accomplies par des facteurs protéiques comme c'est le cas par exemple des microARN.
Structure de l'ARN
Les ribonucléotides
Structure chimique

L'ARN est un acide nucléique, c'est-à-dire une molécule constituée d'un enchaînement de nucléotides. Chaque nucléotide de l'ARN est constitué d'un pentose, le ribose, dont les atomes de carbone sont numérotés de 1' à 5', d'une base azotée et d'un groupement phosphate. La base azotée est reliée par un atome d'azote au carbone 1' du ribose. Les nucléotides sont liés les uns aux autres par des groupements phosphate, par l'intermédiaire de liaisons phosphodiester au niveau des carbones 3' et 5'. L'ARN possède quatre bases azotées différentes : l'adénine (notée A), l'uracile (noté U), la cytosine (notée C) et la guanine (notée G). La thymine de l'ADN est remplacée par l'uracile dans l'ARN. La différence entre ces deux bases est le remplacement d'un groupement méthyle en position 5 de la thymine par un hydrogène dans l'uracile. Cette modification de structure ne change pas les propriétés d'appariement avec l'adénine.
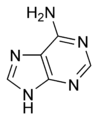 Adénine. |  Guanine. | 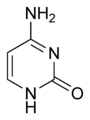 Cytosine. | 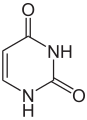 Uracile. |
Stéréochimie


Sur le plan structural, la présence d'un oxygène sur la position 2' du ribose influence la conformation du cycle furanose du ribose. Ce cycle à cinq atomes n'est pas plan, ce qui conduit à deux conformères principaux du sucre, appelés C2'-endo ou C3'-endo. Dans l'ARN qui comporte un oxygène en position 2', la position C3'-endo est privilégiée, ce qui modifie profondément la structure des doubles hélices comportant des brins ARN. Ces duplex d'ARN forment une hélice de type A, différente de celle qui est observée de façon majoritaire dans l'ADN classique qui est une hélice de type B, où le désoxyribose est en conformation C2'-endo.
Double hélice d'ARN

L'hélice de type A qu'adopte l'ARN lorsqu'il forme un duplex, a des propriétés géométriques assez différentes de celles de l'hélice de type B. Tout d'abord le nombre de paires de bases par tour d'hélice est de 11 au lieu de 10 pour l'ADN en forme B. Le plan des paires de bases n'est plus perpendiculaire à l'axe de l'hélice, mais forme un angle d'environ 75 degrés avec celui-ci. Il en résulte un déplacement de l'axe de l'hélice qui ne passe plus par le centre de l'appariement des bases, mais à l'intérieur du grand sillon. Ceci induit une augmentation du diamètre de l'hélice qui passe d'environ 20 Å pour l'ADN en forme B à environ 26 Å pour l'ARN en forme A. Enfin, la géométrie des deux sillons est profondément affectée : le petit sillon devient très accessible, tandis que le grand sillon devient très profond, étroit et pincé. Ceci a un impact sur la manière dont l'ARN en duplex apparié peut interagir avec des protéines, car l'étroitesse du grand sillon est une barrière à l'accessibilité de ligands protéiques.
Structure in vivo
La plupart des ARN naturels sont présents sous forme simple brin dans la cellule, contrairement à l'ADN qui est sous forme d'un double-brin apparié. Les ARN simple brin se replient le plus souvent sur eux-mêmes, formant une structure intramoléculaire qui peut être très stable et très compacte. La base de cette structure est la formation d'appariements internes, entre bases complémentaires (A avec U, G avec C et, parfois, G avec U). La description des appariements internes entre les bases d'un ARN s'appelle la structure secondaire. Cette structure secondaire peut être complétée par des interactions à longue distance qui définissent alors une structure tridimensionnelle ou structure tertiaire.
La formation de la structure des ARN est très souvent dépendante des conditions physico-chimiques environnantes et en particulier de la présence de cations divalents, comme le magnésium Mg2+, dans la solution. Ces cations interagissent avec les groupements phosphate du squelette et stabilisent la structure, en particulier en faisant écran à la répulsion électrostatique entre les charges négatives de ces phosphates.
L'existence de structure tertiaire dans les ARN est à la base de la richesse de ses fonctions et en particulier de sa capacité à catalyser des réactions chimiques (ribozymes).
Structure secondaire

La structure secondaire d'un ARN est la description de l'ensemble des appariements internes au sein d'une molécule simple brin. Cet ensemble d'appariements induit une topologie particulière, composée de régions en hélice (tiges) et de régions non-appariées (boucles). Par extension, la structure secondaire recouvre également la description de cette topologie.
L'élément moteur de la formation de structures secondaires au sein d'un ARN simple brin est l'existence de régions contenant des séquences répétées inversées, qui peuvent s'apparier pour former localement une structure en double hélice. Par exemple, si l'ARN contient les deux séquences suivantes : --GUGCCACG----CGUGGCAC--, celles-ci forment un motif répété inversé, les nucléotides du second segment étant les complémentaires de ceux du premier, après inversion de leur sens de lecture. Ces deux segments peuvent donc s'apparier de manière antiparallèle pour former une région localement en duplex. La région entre les deux segments forme alors une boucle reliant les deux brins du duplex. On parle alors de structure en tige et boucle ou en « épingle à cheveux ».

Dans des ARN de longueur plus importante, il peut exister des structures plus complexes qui résultent de l'appariement de plusieurs régions complémentaires ou séquences répétées inversées. En fonction de la manière dont sont « emboîtées » ces différentes régions, on obtient des éléments topologiques variés, avec des tiges ou régions appariées, et divers types de boucles :
- les boucles terminales, situées à l'extrémité d'une tige ;
- les boucles internes, qui connectent deux tiges ;
- les boucles multiples, qui connectent trois tiges ou plus et constituent des points de branchement de la structure ;
- les hernies (en anglais bulge), ou boucles latérales, qui sont sur un seul des deux brins d'une hélice. La continuité de l'hélice n'est en général pas affectée et les bases restent empilées de manière coaxiale, de part et d'autre de la hernie.
Il n'existe pas toujours une structure unique stable pour une séquence donnée et il arrive que certains ARN puissent adopter plusieurs conformations alternatives en fonction de la liaison d'un ligand (protéine, petite molécule…) ou des conditions physico-chimiques (force ionique, pH). On peut en général suivre la formation ou la fusion de la structure secondaire d'un ARN par des mesures spectroscopiques. Ainsi, par exemple, l'absorption dans l'ultraviolet des bases de l'ARN est plus importante à l'état déplié qu'à l'état replié (phénomène d'hyperchromicité).
Structure tertiaire

Appariements non canoniques
Au-delà de la topologie des boucles et des hélices composées de paires de bases standard, un ARN peut adopter une structure tridimensionnelle compacte, ou structure tertiaire, comme une protéine. À l'intérieur de cette structure, les hélices canoniques sont complétées par des appariements non canoniques, c'est-à-dire distincts des appariements classiques, de type Watson-Crick (A-U et G-C) et bancals (wobble, G-U). On a observé une grande variété de ces appariements dans les structures tridimensionnelles d'ARN résolues par cristallographie ou par résonance magnétique nucléaire. On trouve par exemple des appariements Hoogsteen et des appariements « en cisaille » (sheared). Il existe également des interactions base-ribose, notamment avec le 2'-hydroxyle, qui peut former des liaisons hydrogènes. Une nomenclature systématique de toutes ces interactions a été proposée par Eric Westhof et ses collaborateurs. Plus de 150 types d'appariements ont été observés et ont été regroupés en douze grandes familles. Ces appariements non canoniques impliquent toujours des liaisons hydrogènes entre les bases, qui sont coplanaires, comme dans les paires Watson-Crick.
Interactions à longue distance
Des appariements canoniques ou non canoniques peuvent intervenir entre des régions distantes de la structure secondaire, souvent localisées dans des boucles, ce qui permet de stabiliser un repliement compact de la structure.
Parmi ces interactions non canoniques à longue distance figurent :
- les pseudonœuds, structures formées par l'interaction d'une boucle avec une région située à l'extérieur de la tige qui la délimite ;
- les triplex de brin, qui surviennent lorsqu'une région simple brin vient s'insérer dans le grand sillon d'une région en hélice ;
- les interactions tétraboucle-récepteur : interactions entre boucles hyperstables de quatre nucléotides (tétraboucles) et structures en duplex ou quasi-duplex.
Similitudes et différences entre l'ADN et ARN
Les principales différences entre les deux molécules sont que :
- l'ARN a pour composant un sucre ribose là où l'ADN a un désoxyribose ;
- la base uracile a un rôle pour l'ARN qui est rempli par la thymine dans l'ADN ;
- l'ARN existe généralement sous la forme d'un simple brin (monocaténaire), sauf chez quelques virus comme les reovirus, tandis que l'ADN est double brin (bicaténaire) avec une structure en double hélice ;
- l'ARN est court : de quelques dizaines à quelques milliers de nucléotides pour les ARN cellulaires (ARNm ou ARN structurés), contre quelques millions à quelques centaines de millions dans l'ADN qui constitue le génome de la cellule.
Les trois premières différences donnent à l'ARN une stabilité bien moindre que celle de l'ADN :
- le ribose possède un groupement hydroxyle en position 2', qui est absent dans le désoxyribose de l'ADN. Cette fonction 2'-OH a des incidences multiples sur la structure de l'ARN. Tout d'abord sur le plan chimique, cette fonction alcool rend l'ARN sensible à l'hydrolyse alcaline. La présence des deux oxygènes en cis sur les positions 2' et 3' rend possible la cyclisation du phosphate sur les positions 2' et 3', qui se produit très rapidement lorsqu'une base vient arracher le proton du 2'-OH. Cette cyclisation du nucléotide provoque une coupure de la chaîne ribose-phosphate et libère des extrémités 5'-OH et 2', 3' phosphate cyclique ;

- l'uracile est moins coûteux énergétiquement à produire pour les organismes vivants que la thymine, puisqu'il nécessite une étape de synthèse de moins, qu'est la méthylation par la thymidylate synthase. La présence de thymine dans l'ADN permet à la cellule de détecter des lésions spontanées de la cytosine qui est sensible à l'oxydation. La désamination spontanée de la cytosine en présence d'oxygène convertit cette dernière en uracile. La présence de désoxyuracile dans l'ADN est anormale, puisque le désoxyribonucléotide complémentaire du A est la thymidine. Grâce à cette distinction thymine/uracile, la machinerie de réparation par excision de base peut détecter et corriger le défaut. Dans l'ARN, la désamination des cytosines produit des uraciles et n'est pas réparée. L'ARN a une durée de vie beaucoup plus courte que celle de l'ADN, il est dégradé et recyclé ;
- si un brin d'ARN monocaténaire est endommagé, la lésion n'est pas réparée et le dommage est irréversible ; en revanche, si un des deux brins d'ADN est endommagé, la cellule peut utiliser l'information portée brin par le brin complémentaire intact pour réparer la lésion.
D'un point de vue évolutif, certains éléments permettent de penser que l'ARN serait antérieur à l'ADN comme support de l'information génétique, ce qui expliquerait ses fonctions plus étendues et sa généralisation. L'ADN serait apparu plus tard et n'aurait supplanté l'ARN que pour le rôle de stockage à long terme, en raison de sa plus grande stabilité.